Notes
The following text explores, in a speculative manner, the evolutionary question: Did high-intensity affective states, specifically Pain, emerge early in evolutionary history, or did they develop gradually over time?
Note: We are not neuroscientists; our work draws on our evolutionary biology background and our efforts to develop welfare metrics that accurately reflect reality and effectively reduce suffering. We hope these ideas may interest researchers in neuroscience, comparative cognition, and animal welfare science.
This discussion is part of a broader manuscript in progress, focusing on interspecific comparisons of affective capacities—a critical question for advancing animal welfare science and estimating the Welfare Footprint of animal-sourced products.
Key points
Ultimate question: Do primitive sentient organisms experience extreme pain intensities, or fine-grained pain intensity discrimination, or both?
Scientific framing: Pain functions as a biological signalling system that guides behavior by encoding motivational importance. The evolution of Pain signalling —its intensity range and resolution (i.e., the granularity with which differences in Pain intensity can be perceived)— can be viewed as an optimization problem, where neural architectures must balance computational efficiency, survival-driven signal prioritization, and adaptive flexibility.
Mathematical clarification: Resolution is a fundamental requirement for encoding and processing information. Pain varies not only in overall intensity but also in granularity—how finely intensity levels can be distinguished.
Hypothetical Evolutionary Pathways: by analysing affective intensity (low, high) and resolution (low, high) as independent dimensions, we describe four illustrative evolutionary scenarios that provide a structured framework to examine whether primitive sentient organisms can experience Pain of high intensity, nuanced affective intensities, both, or neither.
Introduction
Humans possess a rich affective spectrum, where pain and pleasure range from mild sensations to extreme states like excruciating pain or overwhelming bliss. Yet, the evolutionary origins of affective experiences still remain a mystery (Schukraft 2020; Birch 2024; Solms 2021; Panksepp et al. 2017; Godfrey-Smith 2016; Feinberg 2024). Aligned with scholars like Panksepp (1998) and Solms (2021), who assert that sentience (or consciousness) itself is defined by the very capacity for valenced (positive or negative) mental experience, here we connect the capacity for affective experiences intrinsically with the emergence of sentience, bypassing the debate over whether sentient organisms experience affective states. Our focus instead centers on a question crucial for prioritizing efforts to reduce suffering: when, in evolutionary history, the capacity to experience negative affective states (‘Pain’, for short) of high intensity emerged. Specifically, we explore whether sentient organisms with simpler nervous systems— such as annelids (e.g., earthworms) or arthropods (e.g., insects, crustaceans)—can experience Pain reaching 'Disabling' or 'Excruciating' levels[1] (see Alonso & Schuck-Paim, 2021 or this source for definitions).
To explore whether high-intensity Pain can be experienced by primitive sentient organisms, we reframe the question as one of information resolution within a scale of Pain intensities. Briefly, two evolutionary possibilities are considered for how nervous systems evolved to represent varying Pain intensities, enabling appropriate responses to competing behavioural demands: (1) increasing resolution within a fixed range of intensities (i.e., introducing finer gradations between the minimum and maximum perceived intensity, e.g., 0 to 10) or (2) expanding the range itself of perceived intensities (e.g., extending the maximum perceived intensity value beyond 10, to values like 100, or 1000). These can be understood as alternative (non-exclusive) evolutionary solutions for encoding motivational salience in nervous systems—that is, for signaling the urgency or importance of a stimulus in order to prioritize behavioral responses that enhance survival. Each strategy would likely leave distinct neurobiological signatures, offering a tractable way to study their evolution.
The Function and Evolution of Affective Scales
From a quantitative standpoint, any system that encodes and processes information—whether sensory perception, motor control, or affective states—requires a scale with a certain resolution (Shannon 1948). This principle applies broadly across disciplines, from signal processing in engineering to perception in biological systems.
Pain and pleasure serve as an adaptive motivational signal, guiding organisms toward behaviors that enhance survival and reproductive success. A scale of multiple intensities allows organisms to prioritize responses based on urgency: high-intensity pain triggers immediate, drastic action (e.g., fleeing from a predator, attending to an urgent biological need such as water or oxygen) overriding other behaviours, whereas a more moderate pain intensity allows for subtler behavioral adjustments (e.g., shifting posture to relieve pressure).
From a human perspective, it seems intuitive to view pain as a spectrum from 'No Pain' to an unbearable 'Excruciating' level. However, like many biological adaptations, the human pain scale may represent just one of several evolutionary solutions for eliciting behavioral responses to varying threats and needs[2] [^2].
The structure of an affective scale can vary significantly across species and may be shaped by two key dimensions (Figure 1):
Range: The maximum ‘absolute’ value of an affective experience that an organism can perceive, defining the absolute amplitude between the highest or lowest affective intensities possible. A high-range system allows for extreme Pain or pleasure, which may be necessary for strong motivational reinforcement, while a low-range system limits organisms to milder affective states.
Resolution: How precisely an organism can differentiate between varying levels of affective intensity. A high-resolution system allows for fine distinctions, while a low-resolution system may encode only broad categories, limiting the precision of affective experiences[3].
For simplicity, we will henceforth refer to 'Range of Intensity' and 'Resolution of Intensity' as simply 'Range' and 'Resolution,' respectively.
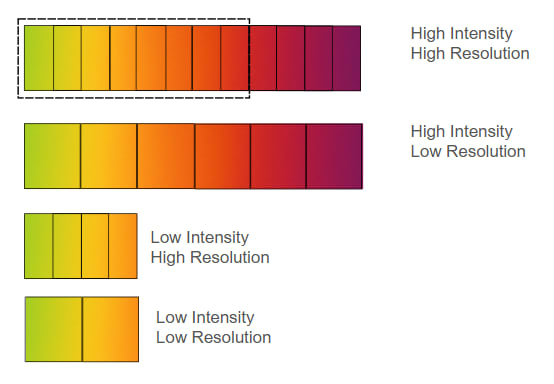
Figure 1. Four configurations of Pain systems in sentient organisms, varying by intensity (range from No Pain to Excruciating, shown as a green-to-purple gradient) and resolution (discrimination ability, shown as segment count: more segments = higher resolution)
From a pure information-enconding perspective, resolution is as versatile as range for enabling intensity-based prioritization of behavioral responses, since in theory both could be theoretically increased indefinitely (range can increase to infinity and resolution can be increasing through an infinite number of subdivisions). However, this theoretical potential is constrained by the realities of biological systems. For example, the resolution of a sensory system, such as the eye, is limited by the size and spacing of the photoreceptor cells that detect light, limiting the resolution of visual perception. Affective states are likely to similarly face neurological and energetic trade-offs that limit the "bit depth" (i.e., the number of discriminable affective intensities) that an organism can encode and process within a given intensity range.
In that sense, it is reasonable to expect that brains with greater neuronal complexity will tend to have a larger number of potential discriminable states, that is, greater affective intensity resolution. This follows from fundamental information processing principles - a system with more components and connections can encode more distinct states, just as a digital system with more bits can represent more unique values, allowing for finer gradations along a continuous scale of intensity. This does not necessarily mean that simpler nervous systems experience lower maximum affective intensities, but rather that they likely distinguish between fewer discrete levels within their intensity range. This information-processing capacity, however, is likely to evolve in response to ecological pressures. The decision needs or constraints of organisms greatly vary. Flies, for instance, are likely to have fewer nuances in their behavioral choices (e.g., predator escape, initiate food seeking, posture adjustment); therefore, an affective scale with a lower number of states may suffice.
Which is evolutionary cheaper for a Pain scale: high resolution or wide range?
To address whether primitive sentient organisms can experience extreme Pain intensities, we first consider the evolutionary costs of developing either higher levels of Pain intensity or finer-grained intensity discrimination.
Costs of Increasing Range
Generating higher-intensity affective states likely involves greater activation within nociceptive pathways and broader engagement of central processing systems. In both vertebrates and invertebrates, increasing signal strength is associated with recruiting additional processing units and enhanced synaptic transmission, which demand greater energy (Coghill et al. 1999; Hu et al. 2020)(Yuan, Huo, and Fang 2018). Given these energetic demands, primitive organisms with simpler neural architectures might have evolved Pain systems with intensity ranges tailored to their behavioral repertoires—intense enough to override competing motivations when necessary, but not extending to extreme intensities unless required by their ecological needs and behavioral options.
Costs of Increasing Resolution
In sensory and cognitive systems, increased resolution is generally associated with expanded neural representation, involving either a higher number of processing units or more complex patterns of connectivity. This pattern holds across modalities, from vision to proprioception. In affective systems, encoding multiple distinguishable levels of pain or discomfort would likely require circuits capable of maintaining and comparing activation thresholds, differentiating between intensities based on stimulus duration, magnitude, or context, and interacting with motivational systems to appropriately modulate behavior.
Like with intensity range, the metabolic and developmental costs associated with building and maintaining the neural architecture to support greater resolution are non-trivial. Evolutionary gains from fine-grained affective discrimination would thus be expected only in contexts where subtle variations in internal state carry important consequences for fitness.
In summary, while limited resolution may evolve under relatively low energetic and structural demands, the development of high-resolution affective systems likely incurs significant neurobiological costs. As a result, high affective resolution is unlikely to be present in primitive sentient organisms unless it serves a function that could not be achieved through simpler, lower-cost alternatives.
Trajectories for the Evolution of Range and Resolution
Building on the trade-offs between range and resolution, we propose four hypothetical scenarios illustrated in Figure 2 that reflect how Pain scales might have emerged in primitive sentient organisms. The human-like ‘high intensity, high resolution’ (HiHr) state is marked by a circle in Figure 2, illustrating a scenario where this configuration may have been present early in the evolutionary history of animals. In contrast, the three alternative scenarios (LiLr, HiLr, and LiHr) shown in the figure indicate potential evolutionary transitions from differing initial states.
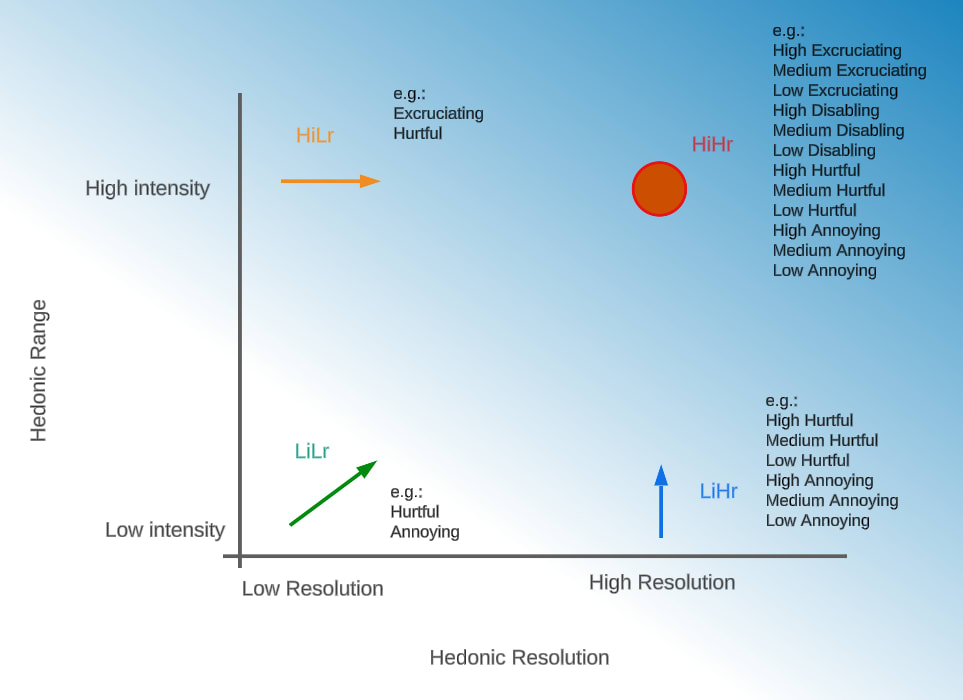
Figure 2. Hypothetical Evolutionary Pathways for Pain intensity evolution, mapped across two dimensions: Hedonic Resolution, representing the ability to distinguish fine gradations of intensity, and Hedonic Intensity Range. LiLr (Low Intensity, Low Resolution): Limited discrimination and experiential intensity range ; HiLr (High Intensity, Low Resolution): Intense but undifferentiated affective states; LiHr (Low Intensity, High Resolution): Mild but nuanced experiential intensities; HiHr (High Intensity, High Resolution): Intense and differentiated experiences . A light-to-dark blue gradient reflects increasing neurological complexity. Each quadrant includes examples of intensities that are possibly felt in each case, using the Welfare Footprint Framework intensity categories. Since the human HiHr state is the only one known to exist with certainty (in humans), it is marked by a red circle, suggesting a hypothesis where this configuration may have persisted from early sentient organisms to humans without major changes. In contrast, LiLr, HiLr, and LiHr (with green, orange, and blue arrows, respectively) imply possible evolutionary transitions from differing initial states.
Low Resolution, Low Intensity (LrLi): Basic Survival Signals
In this scenario, primitive sentient organisms emerged with low resolution and low intensity Pain systems, limited to coarse, survival-driven subjective signals. Their rudimentary nervous systems, such as simple ganglia or decentralized neural architectures, likely lack the neural complexity and metabolic capacity to encode fine gradations or sustain strong subjective Pain. A system with a narrow affective range and few discriminable states could still fulfill adaptive functions (e.g., no Pain, Annoying, or Hurtful Pain) to guide behaviors critical for survival, such as avoiding noxious stimuli, and seeking food. Even extreme survival behaviors could be triggered by weak signals, as computational efficiency can allow strong reactions from minimal inputs. This hypothesis aligns with proposals that primitive sentient organisms prioritize efficiency over richness in subjective experience (Sterling & Laughlin, 2015).
High Resolution, Low Intensity Range (LiHr): Subtle but Mild Signals
In this “cheap high-resolution” scenario, primitive organisms would already have a complex repertoire of states within a low range of intensities. Since moderate gains in resolution may be achieved through localized circuit elaboration without systemic amplification, this trajectory is energetically plausible for organisms with increasingly complex neural architectures. In such cases, increases in intensity range would have emerged later in evolutionary history (e.g., if further increases in resolution were no longer possible, or if more complex structures enabled greater signal amplification).
Low Resolution, High Intensity (HiLr): Strong but Undifferentiated Signals
In this “cheap high-intensity” scenario, early sentient organisms would have already evolved an aversive response system with strong but undifferentiated reactions to critical survival threats and biological needs. Rather than distinguishing between subtle variations in discomfort, these organisms would encode pain or pleasure as broad, overpowering signals.
This scenario assumes that high affective intensities can emerge on simple neural circuits, or alternatively that early sentient organisms have robust signal amplification mechanisms—such as increased neuronal firing rates and widespread neuromodulator release—to produce intense but coarse affective states (LeDoux, 2015), without specialized circuits for fine-grained discrimination. However, this path likely entails significant physiological costs, including mobilizing high-energy stress responses and maintaining systemic readiness for strong affective outputs.
A theoretical extreme of the HiLr model would be an organism with a binary pain system—experiencing either No Pain or the worst possible Pain (Tomasik 2019). While intriguing, this idea treats affective perception as a purely logical construct, ignoring the neural and physiological constraints on pain processing. Even in abstract terms, it suggests that primitive organisms would experience near-infinite suffering, decreasing only as neural complexity increased. However, if affective intensity is subject to the thermodynamic and biological limitations discussed, this extreme seems implausible.
High Resolution, High Intensity (HiHr): Rich and Extreme Signals
This scenario proposes that the full spectrum of rich, nuanced, and extreme Pain experiences observed in modern vertebrates and cognitively sophisticated species was not evolutionarily or physiologically costly, making it possible for even primitive sentient organisms to experience both high and nuanced affective intensities.
For this capacity to exist in more primitive organisms, it would be necessary to assume that there would be no true “primitive” stage in terms of affective valence—soon after sentience emerged, it would have done so with a nearly fully developed capacity for extreme pain and pleasure, rather than evolving gradually from lower intensities. This could be explained by a neurological threshold effect: if the minimum cognitive complexity required for sentience is already high, then organisms crossing that threshold might inherently possess the neural architecture needed to support the full spectrum of motivational salience and hedonic scale.
Tentative Conclusion
Sentience likely emerged independently across multiple phylogenetic lineages, minimally including vertebrates, coleoid cephalopods (e.g., octopuses, squids, cuttlefish), arthropods (e.g., insects, decapods like crabs), and possibly onychophorans (velvet worms) (Feinberg & Mallatt, 2016; Ginsburg & Jablonka, 2019; (Feinberg 2024). The core question explored here is whether primitive sentient organisms—such as annelids or certain arthropods—are capable of experiencing pain at intensities comparable to those seen in cognitively complex vertebrates. Specifically, can their subjective states reach levels of suffering that would be classified as Disabling or Excruciating in humans?
To explore this, we developed a framework distinguishing two dimensions of affective architecture: Range (the amplitude of the affective signal), and Resolution (the granularity with which that signal can be represented and behaviorally modulated). This separation allows us to model multiple evolutionary trajectories, , each giving rise to a set of plausible affective configurations.
Those configurations may support adaptive function, but they differ significantly in welfare implications. Those that depart from low intensity ranges, (LiLr) and LiHr), suggest that primitive sentient organisms, including those extant today, do not experience concerning Pain levels. By contrast, pathways that depart from high intensities (HiLr and HiHr) imply that even primitive sentient organisms could experience extremely intense Pain, expanding extraordinarily the range of organisms capable of suffering at morally concerning levels. Distinguishing between those pathways requires methods focused on the presence of resolution and on the magnitude of affective intensity.
To further investigate these hypotheses, we propose two empirical approaches:
Behavioral complexity analysis: If primitive sentient organisms have a higher-resolution Pain system, their behavioral repertoire should reflect graded responses.
- Testing Approach: By analyzing the degrees of behavioral freedom and emergency response strategies available to primitive sentient organisms, we can estimate the minimum resolution of their affective scales. This could involve quantifying behavioral variability in response to noxious stimuli and comparing it across species with known differences in neural complexity.
- Important Clarification: Higher intensity can sometimes act as a proxy for higher resolution, since a wider range of possible intensities can create finer distinctions in behavioral output. However, intensity alone does not explain how organisms modulate their responses within that range—which is the key question when distinguishing resolution from raw intensity.
Neuroenergetic assessment: If primitive organisms experienced high-intensity pain, we would expect to see neurobiological adaptations that support extreme affective states—such as highly active nociceptive pathways, complex neuromodulatory mechanisms, and energy-intensive pain-processing circuits. If they instead evolved low-intensity pain with high resolution, we would expect a greater investment in neural structures that refine sensory discrimination, allowing for fine-grained affective states without necessarily reaching extreme intensities.
- Testing Approach: Measuring the energetic and computational demands of pain-processing systems across species could help determine whether primitive nervous systems had the capacity to support high-intensity pain. This could involve functional neuroimaging, metabolic studies, or neural activity mapping to examine whether these organisms allocate enough energy to support highly intense or highly differentiated pain experiences.
- Important Clarification: While higher intensity pain may require more neural energy, the same is true for higher resolution, since finer discrimination among affective states demands more complex neural computations. Therefore, this analysis should control for both factors—distinguishing between species that evolved higher affective resolution without greater intensity versus those that evolved stronger pain signals without increasing resolution.
These investigations have significance beyond their contribution to evolutionary biology, carrying profound implications for animal welfare ethics, particularly regarding the use of invertebrates in agriculture, research, and other human activities. If primitive sentient organisms experience only low- to moderate-intensity Pain (e.g., LiLr and LiHr), high suffering capacity might have appeared later in evolution at undetermined points. Conversely, if primitive nervous systems could generate extreme intensities (HiLr and HiHr), this expands the range of organisms capable of morally concerning Pain levels, broadening our ethical obligations toward them (Birch, 2017; Browning, 2019).
Acknowledgements
We are grateful to Vasco Grilo for his insightful comments on an earlier draft of this text.
References
Alonso, W. J., & Schuck-Paim, C. (2021). Pain-Track: A Time-Series Approach for the Description and Analysis of the Burden of Pain. BMC Research Notes, 14(1), 229.
Birch, Jonathan. 2024. The Edge of Sentience: Risk and Precaution in Humans, Other Animals, and AI. London, England: Oxford University Press.
Coghill, R. C., C. N. Sang, J. M. Maisog, and M. J. Iadarola. 1999. “Pain Intensity Processing within the Human Brain: A Bilateral, Distributed Mechanism.” Journal of Neurophysiology 82 (4): 1934–43.
Feinberg, Todd E. 2024. From Sensing to Sentience. London, England: MIT Press.
Godfrey-Smith, Peter. 2016. Other Minds: The Octopus and The Evolution of Intelligent Life. HarperCollins Publishers Limited.
Hu, Yujia, Congchao Wang, Limin Yang, Geng Pan, Hao Liu, Guoqiang Yu, and Bing Ye. 2020. “A Neural Basis for Categorizing Sensory Stimuli to Enhance Decision Accuracy.” Current Biology: CB 30 (24): 4896–4909.e6.
LeDoux, J. (2015). Anxious: Using the Brain to Understand and Treat Fear and Anxiety. Viking.
Panksepp, Jaak, Richard D. Lane, Mark Solms, and Ryan Smith. 2017. “Reconciling Cognitive and Affective Neuroscience Perspectives on the Brain Basis of Emotional Experience.” Neuroscience and Biobehavioral Reviews 76 (Pt B): 187–215.
Schukraft, Jason. 2020. “Differences in the Intensity of Valenced Experience across Species.” Rethink Priorities. October 29, 2020.
Shannon, C. E. 1948. “A Mathematical Theory of Communication.” The Bell System Technical Journal 27 (3): 379–423.
Solms, Mark. 2021. The Hidden Spring: A Journey to the Source of Consciousness. W. W. Norton & Company.
Tomasik, Brian. 2019. “Is Brain Size Morally Relevant?” Essays on Reducing Suffering. 2019. .
Yuan, Ye, Hong Huo, and Tao Fang. 2018. “Effects of Metabolic Energy on Synaptic Transmission and Dendritic Integration in Pyramidal Neurons.” Frontiers in Computational Neuroscience 12 (September):79.
- ^
In the Welfare Footprint framework, pain intensities are defined as absolute measures, meaning that one hour of Excruciating pain in humans is assumed to be hedonically equivalent to one hour of Excruciating pain in shrimps, if shrimps were capable of experiencing Excruciating pain. This approach treats pain intensities as fixed, anchoring them to human experience rather than scaling them based on a species’ welfare range, to ensure comparability across species. This means that (i) If a primitive organism can only experience pain comparable to Annoying in humans, we would ideally classify it as Annoying—not Excruciating, even if it is the worst pain the organism can feel, (ii) if an organism could experience pain exceeding the human Excruciating threshold, the scale would need to be expanded (e.g., “Astronomical” or “Unfathomable”). We acknowledge that this absolute framework presents challenges in the absence of knowledge about the hedonic capacities of each species. This is relevant in practical assessments; for example, in estimating the Cumulative Pain of boiling a lobster alive, knowing whether a lobster’s maximum possible pain intensity is closer to the ‘human’ Annoying, Hurtful, or Excruciating pain is crucial. Since we currently lack knowledge on interspecific differences in hedonic capacity, we provisionally allow for the possibility that Excruciating pain is experienced in cases like the previous example. However, we hold this assumption as temporary until better evidence allows for a more accurate placement of each experience on an absolute scale. Both relative and absolute scaling approaches are valid depending on context, but they should not be mixed without adjustment to avoid double counting or misinterpretation of intensity levels across species.
- ^
Reflex mechanisms, for instance, are earlier evolutionary solutions to the same challenge, triggering responses without requiring subjective experience (Feinberg 2024).
- ^
Resolution is distinct from affective diversity, which refers to the number of qualitatively different types of affective experiences an organism can have (e.g., fear vs. social exclusion vs. hunger). An animal may be capable of experiencing multiple kinds of intense affective states yet possess a coarse resolution system that prevents it from distinguishing degrees of intensity within each type. Conversely, an organism may experience a narrow set of affective types but represent their intensity with high internal resolution. The current framework focuses exclusively on within-modality resolution.
Executive summary: This speculative post explores whether primitive sentient organisms can experience extremely intense pain by disentangling two independent dimensions of affective experience—intensity range and resolution—proposing an evolutionary framework with four possible trajectories and highlighting the ethical importance of determining which organisms might suffer at morally concerning levels.
Key points:
This comment was auto-generated by the EA Forum Team. Feel free to point out issues with this summary by replying to the comment, and contact us if you have feedback.