Hi Vasco. I am new on this forum, so please excuse my lack of knowledge on the specifics of your work over the years. I see that you produce high quantities of seemingly high quality work on undervalued topics such as this one. I myself am very much interested in the sentience and welfare of such “primitive” organisms. Here are a few of my impressions on this post:
Although you take care of summarizing your research and clearly displaying your methodology, your work is still very enigmatic at first glance. Because your conclusion feels counter-intuitive and may spark defensiveness from animal advocates readers, not being able to understand clearly how you came to these conclusions make them feel untrustworthy.
The reasoning that links your work with your conclusion feels… odd. I’m feeling like there are parts missing. For example, from what I understand of this post, you recommend increasing land use to reduce soil animals lifespan because you estimate they have negative lives. However, you thus recommend changes in food consumption towards diets that include more animal-based foods like beef, because they require more agricultural land than any other foods. Why do you consider eating beef to be the most cost-effective option to increase land use ? As I believe most vegans (or vegetarians) developed a strong deontological reasoning for not eating animal-based products, implying that eating beef might be the best utilitarian way to reduce suffering does not feel right: increasing land use is, so why should they tolerate factory farming, or even extensive farming at all ? Are there not any other opportunities to increase land use ? For example, instead of arguing for changes in food consumption, we could argue in favor of considerably increasing the space allocated to farmed animals so that more of the land would be used, farmed animals’ welfare would be increased; and because land is limited, the actual consumption of animal-based products by humans could actually decrease; human health would thus be improved as well. You even considered the possibility of “paving all the Earth’s land”, but nothing in-between. Here’s another scenario: increasing the consumption of beef and land use implies huge greenhouse gas emissions, water pollution, etc -> climate change increases the probability of huge natural disasters and reduces the yield of agricultural production -> billions are spent in damages and health issues that could have been best invested in climate-resilient agricultural practices and wild animals welfare issues. I feel like your conclusion lacks a more systematic view on these issues; and I understand that this work is about utilitarianism, but caring about the welfare of a few species (even in way higher numbers) while recommending more consumption of other poorly-treated species (even in way smaller numbers) feels odd, to me atleast.
I do not get why you consider “targeted research on whether soil animals have positive or negative lives” to be one of the most cost-effective ways of increasing animal welfare in the same way you consider increasing animal-based consumption is, because the latest suggestion implies that the former one already has been made. If research shows that soil animals have positive lives, then your recommendation for more animal-based foods consumption is actually counter-productive, which you acknowledge. I understand that you estimated around 55% probability that they have negative lives, which may be sufficient to consider the horrific possibility that such sheer numbers of animals live horrible lives and work on this question - but it also feels way too close to a 50% chance to actually recommend anything to me. It feels wrong to consider as cost-effective two recommendations that could potentially invalidate each-other: more research would already answer if we should increase land use or not, so recommending eating more beef does not belong here yet.
Your research is very interesting, but I feel like strongly endorsing such concrete and disruptive recommendations (i.e. increasing the consumption of beef) under such uncertainties and hypotheses feels odd. You even argue that eating cheaper plant-based foods allows for more donations, increasing cost-effectiveness even more; which seems to contradict your recommendation. Plus, you mention the “at a margin” problem, as most of the world already eats huge quantities of beef, way above healthy recommendations, so I don’t get why you would recommend eating more beef at all. I feel like such a recommendation would need way more arguments, especially in a systemic analysis of the whole food production chain. That increasing land use could increase global animal welfare is an interesting point - but that we should eat more beef is not a conclusion that I would endorse with this post’s arguments.
Thank you for sharing your work here. I do not expect you to answer thoroughly to every point I made if you do not think your time is worth it - I just felt like sharing a few thoughts on an interesting topic and participating in the forum - but I will gladly read your responses (or anyone's) if you have some.
Great comment and welcome to the forum! looking forward to hearing more of your thoughts :).
Only one small comment which may help understanding @Vasco Grilo🔸 can say if I'm representing him incorrectly. I agree that the uncertainty is so high here that it doesn't make sense to make strong practical recommendations. Vasco though is a mathematical utilitarian in a pretty pure form, so he's seemingly happy to make strong recommendations where there's little evidence and probabilities are close to 50/50. he'll then even change those recommendations immediately after doing some more calculations. I don't really understand how this can work in practice as communities of EA doers obviously can't switch from advocating eating less meat to advocating huge farms on the basis of an extremely uncertain BOTEc. I've made a similar point to you on as few of his posts in the past.
Thanks for the relevant comment, Nick. I strongly upvoted it because I felt it was useful for me to think about this more.
I recommend funding A over B if I think funding A increases welfare more cost-effectively at the margin than funding B (in expectation). However, my numerical estimates of the cost-effectiveness do not integrate all the information I have. If my numerical estimates for the cost-effectiveness of 2 interventions are similar, other factors could easily be decisive.
I agree it would not make sense for a whole community to switch back and forth between 2 interventions with similar positive cost-effectiveness at the margin. The amount of resources moving from the most to the least cost-effective intervention at the margin should tend to 0 as the difference between the marginal cost-effectiveness of the interventions tends to 0.
I also agree it would not make for a single person to be constantly switching back and forth between 2 interventions with similar positive cost-effectiveness at the margin. Spending 1 year to switch to a career path which is 0.1 % more impactful per year, accounting for impact through work and donations, would only make sense if one could work for longer than 1 k years (= 1/10^-3) in the new path. This is way too long, and therefore the change would not be worth it.
However, I think the situation is different in the context of constantly switching back and forth between supporting or not an intervention due to large uncertainty about whether it increases or decreases welfare (in expectation). Constant switching leads to spending resources without achieving nothing, which is worse than supporting positive interventions. However, it is unclear whether achieving nothing is better or worse than supporting an intervention which can easily have a positive or negative cost-effectiveness.
In cases where there is large uncertainty about whether an intervention increases or decreases welfare (in expectation), I believe it is often better to support interventions decreasing that uncertainty. This is a major reason for my top recommendation of decreasing the uncertainty about whether soil nematodes have positive or negative lives, which I consider robustly better than constant switching. As I say in the summary, "I am arguing for, by increasing cost-effectiveness, changes in food consumption which increase agricultural land, the most cost-effective global health interventions, and targeted research on whether soil animals have positive or negative lives".
Thanks for sharing your thoughts, and welcome to the EA Forum, Guillaume!
you recommend increasing land use to reduce soil animals lifespan because you estimate they have negative lives
I recommend decreasing the living time (total animal-years), not lifespan (animal-years per animal), of soil animals via decreasing their density (animals per unit area) given my best guess that they have negative lives.
you thus recommend changes in food consumption towards diets that include more animal-based foods like beef, because they require more agricultural land than any other foods
Right.
implying that eating beef might be the best utilitarian way to reduce suffering does not feel right
I calculate increasing the consumption of beef by 1 kg results in 1.39 billion fewer soil-animal-years, but just in 3.09 more cow-days. In other words, I estimate increasing the consumption of beef decreases the living time of soil animals 164 billion times as much as it increases the living time of cows. I think there is way more suffering in 164 billion soil-animal-years than in 1 cow-year. I calculate increasing the consumption of beef increases the welfare of soil ants, termites, springtails, mites, and nematodes (due to decreasing their living time) 113 k times as much as it changes the welfare of cows for my preferred exponent of the number of neurons of 0.5. The ratio of 113 k is much lower than that of 164 billion because cows have more intense experiences than soil animals. However, increasing beef consumption affects so many more soil-animal-years than cow-years that I still conclude the effects on soil animals clearly dominate.
Why do you consider eating beef to be the most cost-effective option to increase land use ?
I do not. I estimatethat funding HIPF increases agricultural land 25.0 (= 1.29*10^3/51.6) times as cost-effectively as buying beef. As I say in the summary:
"I am arguing for, by increasing cost-effectiveness, changes in food consumption which increase agricultural land, the most cost-effective global health interventions, and targeted research on whether soil animals have positive or negative lives".
For example, instead of arguing for changes in food consumption, we could argue in favor of considerably increasing the space allocated to farmed animals so that more of the land would be used
I think the most cost-effective global health interventions decrease the living time of soil animals much more cost-effectively than interventions aiming to improve the conditions of farmed animals. I estimatethat cage-free and broiler welfare corporate campaigns decrease soil-animal-years 1.13 % (= 5.77*10^7/(5.09*10^9)) and 6.50 % (= 3.31*10^8/(5.09*10^9)) as cost-effectively as funding HIPF.
Here’s another scenario: increasing the consumption of beef and land use implies huge greenhouse gas [GHG] emissions
It is very unclear whether global warming increases or decreases animal welfare. I think it would be valuable to investigate the effects of global warming on soil animals, but this is not my top priority. For a given spending on a land use change intervention (like buying beef, or funding HIPF), I would be surprised if the effects of GHG emissions on soil animals were larger than the direct effects on them resulting from changing their density (the abundance of soil animals per unit area varies by biome).
In any case, the effects of GHG emissions being larger would be a reason for increasing or decreasing (depending on what would benefit soil animals) GHG emissions as cost-effectively as possible. If decreasing GHG emissions was among the most cost-effective ways of increasing the welfare of soil animals, I would recommend Founders Pledge's Climate Fund (FPCF), not interventions targeting farmed animals.
billions are spent in damages and health issues that could have been best invested in climate-resilient agricultural practices and wild animals welfare issues
I do not see how decreasing GHG emissions would be a cost-effective way of fundraising for wild animal welfare. I estimate only 5.03 M 2023-$ were spent on wild animal welfare in 2023. This is a very tiny fraction of the global economy, so decreasing spending in areas outside wild animal welfare would basically have no impact on its funding.
I understand that you estimated around 55% probability that they have negative lives, which may be sufficient to consider the horrific possibility that such sheer numbers of animals live horrible lives and work on this question - but it also feels way too close to a 50% chance to actually recommend anything to me.
Yes, I guess the probability of soil animals having negative lives is around 55 %. Research on whether soil animals have positive or negative lives would be valuable in either case, and this is a major reason I recommend it over funding HIPF.
I prefer sharing my uncertain views about topics I have thought about over being agnostic. There are no interventions outside research which I am reasonably confident increase welfare in expectation. I think electrically stunning shrimp is one of those which more clearly increases welfare in expectation, but I would say it is still unclear whether it increases or decreases welfare in expectation due to effects on soil animals. For my individual welfare per animal-year proportional to "number of neurons"^0.5, I determined electrically stunning shrimp increases the welfare of these by 0.00144 QALY/shrimp. Thereare 94.3 shrimps per shrimp-kg. So infer electrically stunning shrimp increases the welfare of these by 0.136 QALY/shrimp-kg (= 0.00144*94.3). For my individual welfare per animal-year proportional to "number of neurons"^0.5, I estimate replacing farmed shrimp with farmed fish changes the welfare of soil ants, termites, springtails, mites, and nematodes by 364 QALY/shrimp-kg (= 522 - 158). So I conclude electrically stunning farmed shrimp changes the welfare of soil animals more than it increases the welfare of shrimps if it results in the replacement of more than 0.0374 % (= 0.136/364) of the consumption of the affected farmed shrimp by farmed fish. I can easily see this happening for even a slight increase in the cost of shrimp. Moreover, there are 3*10^29 soil archaea and bacteria, 613 M (= 3*10^29/(4.89*10^20)) times as many as soil nematodes (the most abundant soil animals), and I would not be surprised if the effects on soil archaea and bacteria were much larger than those on soil animals, which further contributes to the uncertainty about whether electrically stunning shrimp increases or decreases welfare in expectation.
Side note. Personally, I value decreasing 1 h of suffering as much as creating 1 h of happiness if they have the same intensity. However, many people are suffering-focussed, and therefore value averting 1 h of suffering more than creating 1 h of happiness even if they have the same intensity, and do not want to create additional animal lives even if they are positive. For such suffering-focussed people, it matters less whether soil animals have positive or negative lives. I believe there is a pretty strong case for them wanting to increase agricultural land due to this decreasing animal-years a lot.
You even argue that eating cheaper plant-based foods allows for more donations, increasing cost-effectiveness even more; which seems to contradict your recommendation.
I do not think there is a contradiction. One could work on decreasing GHG emissions while driving a car to the office to save time, and therefore decrease GHG emissions more. Likewise, one could argue for increasing the consumption of beef while not eating beef to be more productive, and save money to donate more.
Plus, you mention the “at a margin” problem, as most of the world already eats huge quantities of beef, way above healthy recommendations, so I don’t get why you would recommend eating more beef at all.
I estimate increasing the consumption of beef by 1 kg results in 9.06 fewer human-minutes due to negatively affecting human health, which would be 1.72*10^-5 (= 9.06/60/24/365.25) fewer QALYs assuming only fully happy life is lost (which slightly overestimates the negative effects on humans). At the same time, I calculate increasing the consumption of beef by 1 kg results in 1.39 billion fewer soil-animal-years, and increases the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes by 19.3 kQALY for my preferred exponent of the number of neurons of 0.5. So I estimate the increase in the welfare of animals is 1.12 billion (= 19.3*10^3/(1.72*10^-5)) times the decrease in the welfare of humans.
I think the focus on beef consumption is better explained by flipping the argument: if you are advocating for a reduction in beef consumption, you may be advocating for a substantial increase in animal suffering, and this should give you pause if the reason you are advocating is that you care deeply and desperately about animal suffering.
The general conclusion being that more research is needed in this area for people who care about soil animal welfare to work out whether soil animals live positive or negative lives, and only once this is done work out how this might be tractably actioned to increase or decrease the number of soil animals via land use change.
I'm honestly really hoping that soil animal lives work out net positive at the mite and springtail level, and that nematodes are sufficiently simple as to not possess the kind of conscious experience needed for welfare capacity. But I remain open to the troubling possibility of well-evidenced alternate conclusions, and support Vasco's work to shed light on this area.
Thanks, Kestrel. I would also prefer soil animals to have positive lives, which would mean a much higher global welfare. However, that also lead to counterintuitive conclusions. I would estimate interventions increasing agricultural land, like cage-free and broiler welfare reforms, and saving human lives, would decrease welfare.
(Vasco asked me to take a look at this post and I am responding here.)
Hi Vasco,
I've been taking a minute to reflect on what I want to say about this kind of project. A few different thoughts, at a few different levels of abstraction.
In the realm of politics, I'm glad the ACLU and FIRE exist, even if I don't agree with them on everything, because I think they're useful poles in the ecosystem. I feel similarly about your work. I think this kind of detailed cost-benefit work on non-standard issues, or on standard issues but that leads to non-standard conclusions, is a healthy contribution to EA, separately from whether I agree with or even understand it.
The main barrier to my engaging deeply with your work is that your analyses hinge on strong assumptions that I have no idea how to verify even in theory. The claim that nematodes live net-negative lives, for instance, which you believe with 55% confidence: I have no clue if this is true. I'm not even sure how many hours I would need to devote to form any belief on this whatsoever. (Hundreds?) In general, I have about 2-3 hours of good thinking per day.
IMO, the top comment on this post expresses the "EA consensus" about your style of analysis -- I notice that it has gotten more upvotes and such than the post itself. One implication of this is that there is some persuasion work to be done get folks onboard with some of your assumptions, stylistic choices, and modes of analysis. Perhaps a post along the lines of "why I write the way I write" (Nietzsche did this) or "The moral philosophical assumptions underpinning my style of analysis" would go some of the way to bridging that gap.
I get the sense that you are building an elaborate intellectual edifice whose many moving parts are distributed in many posts, comments, and external philosophical texts. That's well and good, I also have a "headcanon" about my work and ideas that I haven't fully systematized, e.g. I write almost exclusively about the results of randomized controlled trials without explaining theintellectualfoundations of why I do that. But I think your intellectual foundations are more abstruse and counterintuitive. Readers might enjoy a meta post about those foundations: a "start here to understand Vasco Grilo's writing" primer.
I am generally on board with using the EA forum as an extended job interview, e.g. establishing a reputation as someone who can reason and write clearly about an arbitrary subject. I think you're doing a fine job of that. On the other hand, the interaction with Kevin Xia about whether this work is appropriate for Hive, the downvotes that post received, and the fact that you are the only contributor to the soil animals topic here are face value evidence that writing about this topic as much as you do is not career-optimal. Perhaps it deserves its own forum: soilanimalsmatter.substack.com or something like that? And then you you can actually build up the whole intellectual edifice from foundations upwards. I do this (https://regressiontothemeat.substack.com/) and it is working for me. Just a thought.
Thanks for sharing, Mo! I think it is a pretty good sketch. Some points are a bit outdated, or not quite accurate, but I would say one can definetely get a sense of where I am coming from.
Do you have a sense as to why people haven't quite bridged the inferential gap between wherever they are and your work, despite your (patient, repeated, very thorough) attempts to explain?
With respect to my work on soil animals? I guess people who have read the posts, and like quantitative analyses understand my arguments. However, many of those still disagree on empirical grounds. For example, if they believe the expected total welfare of soil animals is negligible compared with that of farmed animals, in contrast to my estimates. In addition, many disagree for fundamental moral reasons. For instance, if they value averting more intense pain much more strongly than what is justified by its intensity, in contrast to valuing averting pain proportionally to its duration, intensity, and probability as implied by expectationaltotalhedonisticutilitarianism (which I strongly endorse).
By all means, show us the way by doing it better 😃 I'd be happy to read more about where you are coming from, I think your work is interesting and if you are right, it has huge implications for all of us.
That's interesting, but not what I'm suggesting. I'm suggesting something that would, e.g., explain why you tell people to "ignore the signs of my estimates for the total welfare" when you share posts with them. That is a particular style and it says something about whether one should take your work in a literal spirit or not, which falls under the meta category of why you write the way you write; and to my earlier point, you're sharing this suggestion here with me in a comment rather than in the post itself 😃 Finally, the fact that there's a lot of uncertainty about whether wild animals have positive or negative lives is exactly the point I raised about why I have trouble engaging with your work. The meta post I am suggesting, by contrast, motivate and justify this style of reasoning as a whole, rather than providing a particular example of it. The post you've shared is a link in a broader chain. I'm suggesting you zoom out and explain what you like about this chain and why you're building it.
I was not clear in my last comment. I meant my top recommendation of investigating whether soil animals have positive or negative lives does not depend on whether the animal populations I analysed have positive or negative welfare. It depends on interventions changing the welfare of soil animals much more than that of their target beneficiaries in expectation. This is also supported by my estimates that the absolute value of the total welfare of soil animals is much larger than that of other animal populations.
Here is some context about how I make recommendations.
(Caveat: I haven't actually read the post and have not voted on the post. I am inclined to believe that the OP's methodology is generally good given the OP's track record of publishing high-quality posts and comments. It could be the case that this post is an outlier and is actually poorly argued - hence the downvotes)
For those who are downvoting this post, could you please explain why you're downvoting? Off the top of my head, here are some reasons for why people would be downvoting:
You disagree with the conclusions of the post (in which case, it usually makes more sense to disagree-vote)
You disagree with the methodology / think that the methodology is flawed (in which case, a downvote is likely justified)
You feel like the OP "spams" the EA forum with similar arguments and that each new post is not adding value on the margin (in which case, it probably makes sense to downvote?)
If this is how you feel, I'm not sure what the OP should do in response. Should the OP stop posting?
Thanks, Dave. I welcome people to comment about their disagreements with the methodology regardless of their vote. This way I could improve it, or explain why I disagree with their criticisms.
Mild downvote here. The conclusions ("I recommend increasing the consumption of beef") do not follow from the premises ("soil animals have negative lives"), even if true. And the premises are highly uncertain and speculative.
There are perhaps other ways to improve or mitigate soil animal lives, and certainly many other ways to increase agricultural land that do not involve killing and eating cows.
Because of that, it feels like the post is intentionally contrarian for the sake of aggravating others, rather than an earnest attempt to improve the lives of soil animals. I appreciate the detail and raising issues many haven't considered. I'd upvote it if it was framed as raising an issue noting the uncertainty and invitation for consideration, rather than a strong recommendation for something that doesn't follow from the premises.
The conclusions ("I recommend increasing the consumption of beef") do not not follow from the premises ("soil animals have negative lives"), even if true.
I estimate buying beef decreases the living time of soil animals 164 billion times as much as it increases the living time of cows. Do you prefer decreasing the consumption of beef (relative to increasing it) because you believe the experiences of cows are over 164 billion times as intense as those of soil animals? If not, could you elaborate on your reasons for disagreeing with my recommendation?
P1: "This 90 year old is about to send a spam message to 100 million people. That will cause ~2000 years of annoyance and suffering. They have only ~4 years of expected mediocre life left, so it would be better to kill them so they can't send the message."
I see. I think you are implying that I should not recommend increasing the consumption of beef to increase the welfare of soil animals even if these have negative lives because there are other interventions that increase their welfare more cost-effectively. I agree with this last part, but I still think there is value in sharing my belief that increasing the consumption of beef is better than decreasing it. I also discuss better options in the summary:
"I am arguing for, by increasing cost-effectiveness, changes in food consumption which increase agricultural land, the most cost-effective global health interventions, and targeted research on whether soil animals have positive or negative lives".
If you are arguing for increasing agricultural land, there are many other ways to accomplish that. You could promote the use of biofuels. Suggest more people get horses as companion animals. Or many other methods.
Hyper-focusing on eating cows is weird. At this point is seems like a way to self-rationalize that eating cows is not just ok but on net preferable.
Brian Tomasik has a post listing ways of decreasing the living time of invertebrates. Buying beef directly from Brazil is the intervention there for which Brian estimated the highest cost-effectiveness, although Brian only estimated the cost-effectiveness of some. I have followed a plant-based diet for 6 years, and do not miss eating beef. I looked into the cost-effectiveness of buying beef given Brian's early research, and its massive land use requirements relative to other foods.
Your reasoning is certainly interesting to me, and you might even be right.
However, I believe the most problematic point boils down to this: we are being asked to commit an act involving certain, morally grave harm (increasing the suffering of farmed animals, which we know to be sentient) in exchange for a potential benefit (reducing nematode suffering) that rests upon a chain of rather uncertain hypotheses.
Are these organisms sentient?
Are their lives net-negative overall?
What is the moral value of their conscious experience, if they have any?
This idea closely resembles the "fat man" scenario in the trolley problem, only elevated to the nth power. It actively proposes inflicting harm upon someone (the farmed animals / the fat man) for a much greater benefit (the nematodes & Co / the people who would be hit by the train). And this is already an example that many people instinctively reject.
But the situation here is infinitely more paradoxical.
We could frame it this way: "You must push the fat man off the bridge. We don't know if he will stop the train. In fact, we don't even know if there are really people on the tracks to save."
"There is a very low probability that there are billions of people on the tracks (i.e., that nematodes & Co are sentient, that their lives are net-negative, and that their consciousness has a certain moral value)."
"But most, most likely, there is no one there (e.g., they are not sentient; the probability that they are is low, e.g., 6.8\% for nematodes, according to the RP data you cited yourself)."
"Yet, the man you push will surely die."
We are committing a certain murder for a totally speculative benefit. This is a harm that will be 90% or more a large-scale, self-contained harm that yields no benefit whatsoever.
But in reality, an even more fitting analogy might be this, since we are talking about reducing suffering, not saving lives in an absolute sense:
"We know there are several serial killers in the world who will torture a high number of people. For this reason, we take one person, make them be born and raise them, with the sole purpose of constantly torturing them on live TV, so that many of the killers will be distracted by the show and will torture their victims less."
To be truly consistent with what we have discussed, however, it would sound more like this:
"Most probably, there are NO several serial killers in the world who torture many people. But since there is a minimal chance that they do exist, we decide to take a person, make them be born, raise them, and constantly torture them on live TV so that the hypothetical serial killers will be distracted by the show and hypothetically kill their victims earlier."
Now, I know this might be more of an anecdote than a completely rational argument. Since we are forced to act in reality, and we are constantly acting, we must do so with the information available to us, even if it is partial. And that is what you tried to reason about. However, we are essentially exchanging a certain (and grave) harm for an improbable benefit (albeit of astronomical proportions).
And this is without even considering the fact that, for all we know, the lives of these animals might be net-positive. In this case, the harm to the farmed animals would be compounded by the harm to the nematodes & Co. In this scenario, our previous story might sound something like this: "There are no serial killers in the world. We torture a guy on TV thinking there are, and some people who watch the show draw inspiration from it and start killing people themselves."
In general, the uncertainty seems so high as to not justify new actions before obtaining new information that would grant us greater clarity, as you rightly suggest.
Furthermore, given that this involves inflicting harm in exchange for something that could have a massive output, why choose the path of harm when there could be many others? It’s a bit like saying, "Would you push the fat man off the bridge?" "Yes, if it were the only option." Most people would try to find other options, so as not to have to kill the fat man and still save the unfortunates on the tracks. Couldn't we just derail the train with a large rock, for example? Reality is often rich with creative possibilities.
And in this case, I don't see why we should actively inflict harm on someone (the farmed animals) simply because this indirectly achieves something greatly good. Couldn't we just do that good thing ourselves?
Why should we reduce the number of nematodes & Co. through farming, which is harmful in its own right? We might as well kill these animals directly if we decide they must live less! I bet we could be far more efficient. We might as well develop a "killer mixture" for these species, or who knows what else?
Admittedly, I'm venturing into "science fiction" here, and the idea wouldn't be very marketable. But this would be the sensible path.
So, is this a weak hypothesis too? Perhaps we could invent some excuse. It is true that a project openly intending to "kill" nematodes would not succeed, nor could a charity pursuing this mission define itself as such in the eyes of public opinion. But perhaps there could be some ingenious ways to frame this as something good and morally approvable? That way, everyone would not see us as the mad people who want to exterminate nematodes, but as someone doing something good (or perhaps irrelevant). And whose secret purpose is to reduce the number of nematodes on Earth. A win-win situation.
Of course, it is not guaranteed that such a thing is possible, and thus this is also pure speculation, but couldn't finding such a creative solution be a very promising way to do good?
Welcome to the EA Forum, Riccardo! Thanks for the comment. I strongly upvoted it.
There are ways of increasing agricultural land which benefit farmed animals. I estimate cage-free and broiler welfare corporate campaigns, and buying beef increase agricultural-land by 16.4, 93.9, and 51.6 m²-year/$. I think it is safe to say such campaigns increase the welfare of chickens, and I also guess pasture-raised cows have positive lives.
Uncertain effects cannot be neglected if one wants to increase welfare in expectation as I do. Consider these 2 options:
A. 100 % chance of decreasing suffering by 1 h, 10 % chance of decreasing suffering by 5 M hours, and 10 % chance of increasing suffering by 15 M hours. Suffering is increased by 1.00 M hours in expectation (= 1*(-1) + 0.1*(-5 + 15)*10^6).
B. 100 % chance of increasing suffering by 1 h, 10 % chance of increasing suffering by 5 M hours, and 10 % chance of decreasing suffering by 15 M hours. Suffering is decreased by 1.00 M hours in expectation (I just changed the sign of all the effects relative to the scenario above).
Based on the certain effects alone, A is better than B. Decreasing suffering by 1 h with certainty (A) is better than increasing suffering by 1 h with certainty (B). However, accounting for all effects, B is much better than A. Decreasing suffering by 1.00 M hours in expectation (B) is much better than increasing suffering by 1.00 M hours in expectation (A) if one wants to decrease suffering in expectation.
I am now very uncertain about whether increasing agricultural land increases or decreases soil-animal-years. So I now recommend even more strongly investigating the effects on soil animals over increasing animal farming, or funding HIPF. In any case, I agree there are ways of increasing agricultural land much more cost-effectively than through increasing animal farming. "I estimated buying beef is 3.72 % as cost-effective as funding HIPF, and that this decreases 5.07 billion soil-animal-years per $". It is just worth having in mind that the popularity of those interventions matters. I guess it will be very difficult to justify pursuing an intervention based on its effects on soil animals. So identifying the popular interventions which increase the welfare of soil animals the most cost-effectively may be the best one can do.
Thank you Vasco for the welcome and for your prompt reply!
Yes, I understand the underlying logic of your reasoning; my intention was only to highlight how paradoxical it might seem when interpreted from a "narrative" perspective. This is, after all, the way most of us instinctively tend to think/feel, even if it’s often not strictly correct.
Thank you for the update on agricultural land use. I hadn't seen your comment. I am absolutely in agreement with you on that point (regarding the need for more research due to uncertainty).
I was wondering if—in the case that it were true that agricultural land decreases the number of nematodes & Co.—given that we write here so that new ideas might be put into action, and thus based on the consensus these ideas can obtain, wouldn't it be more advantageous to highlight more the non-problematic cases for someone who cares about animals?
For example, for a person deeply committed to the animal cause, it will be harder to accept the solution of purchasing animal products. But confirming the idea that it's better to finance cage-free or broiler chicken campaigns for this additional reason (reducing soil animals) could be much more easily accepted, and thus put into practice. Whereas an idea like consuming more beef might risk provoking repulsion and rejection, perhaps even in a place like this (though this is just a hypothesis, as I don't know this forum that well).
We could call this the "Probability that someone will act upon reading the post." Though I suppose it's difficult to quantify.
Conversely, strongly advocating for a controversial/counter-intuitive thesis, as you have partly done, could also contribute to attracting attention and thus generating the desired effects.
Thinking better about the whole issue, even if nematodes had net-positive lives, the course of action could still be controversial, as it would be practically a perfect example of the Repugnant Conclusion.
In general, you are absolutely right to draw attention to this issue. One could argue that it's probably not a relevant topic. But if it were relevant, it would be extremely relevant. And that fact makes it effectively relevant (at least given our current state of knowledge).
I was wondering if—in the case that it were true that agricultural land decreases the number of nematodes & Co.—given that we write here so that new ideas might be put into action, and thus based on the consensus these ideas can obtain, wouldn't it be more advantageous to highlight more the non-problematic cases for someone who cares about animals?
I would still be very uncertain about whether increasing agricultural land increases or decreases welfare even if I was confident it decreased soil-animal-years. This is because I am very uncertain about whether soil animals have positive or negative lives. However, if increasing agricultural land increased welfare, and the increase in welfare per $ is roughly proportional to the increase in m²-years of agricultural land per $, as I calculated for my preferred exponent of the number of neurons of 0.5, I think it would make sense to advocate for global health interventions over ones targeting farmed animals. There is much more funding going to global health interventions than ones targeting farmed animals, which suggests the fundraising multiplier is higher for the former. Moreover, I estimate global health interventions increase agricultural land more cost-effectively.
In general, you are absolutely right to draw attention to this issue. One could argue that it's probably not a relevant topic. But if it were relevant, it would be extremely relevant. And that fact makes it effectively relevant (at least given our current state of knowledge).
Yes, this is how I think about it. I would not be surprised if the absolute value of the total welfare of soil animals was negligible compared with that of farmed animals. However, based on my current knowledge, I believe the absolute value of the total welfare of soil animals is much larger in expectation than that of farmed animals.
I am arguing for, by increasing cost-effectiveness, changes in food consumption which increase agricultural land, the most cost-effective global health interventions, and targeted research on whether soil animals have positive or negative lives.
I am now very uncertain about whether increasing agricultural land increases or decreases soil-animal-years. I also continue to be very uncertain about whether soil animals have positive or negative lives. So I now just recommend research on the welfare of soil animals.
I estimate the change in the welfare of animals affected by the production of beef, pork, chicken, turkey, dairy milk, fish, eggs, shrimp, peas, tofu, and soy milk. I analyse directly affected animals, and soil ants, termites, springtails, mites, and nematodes impacted by changes in land use.
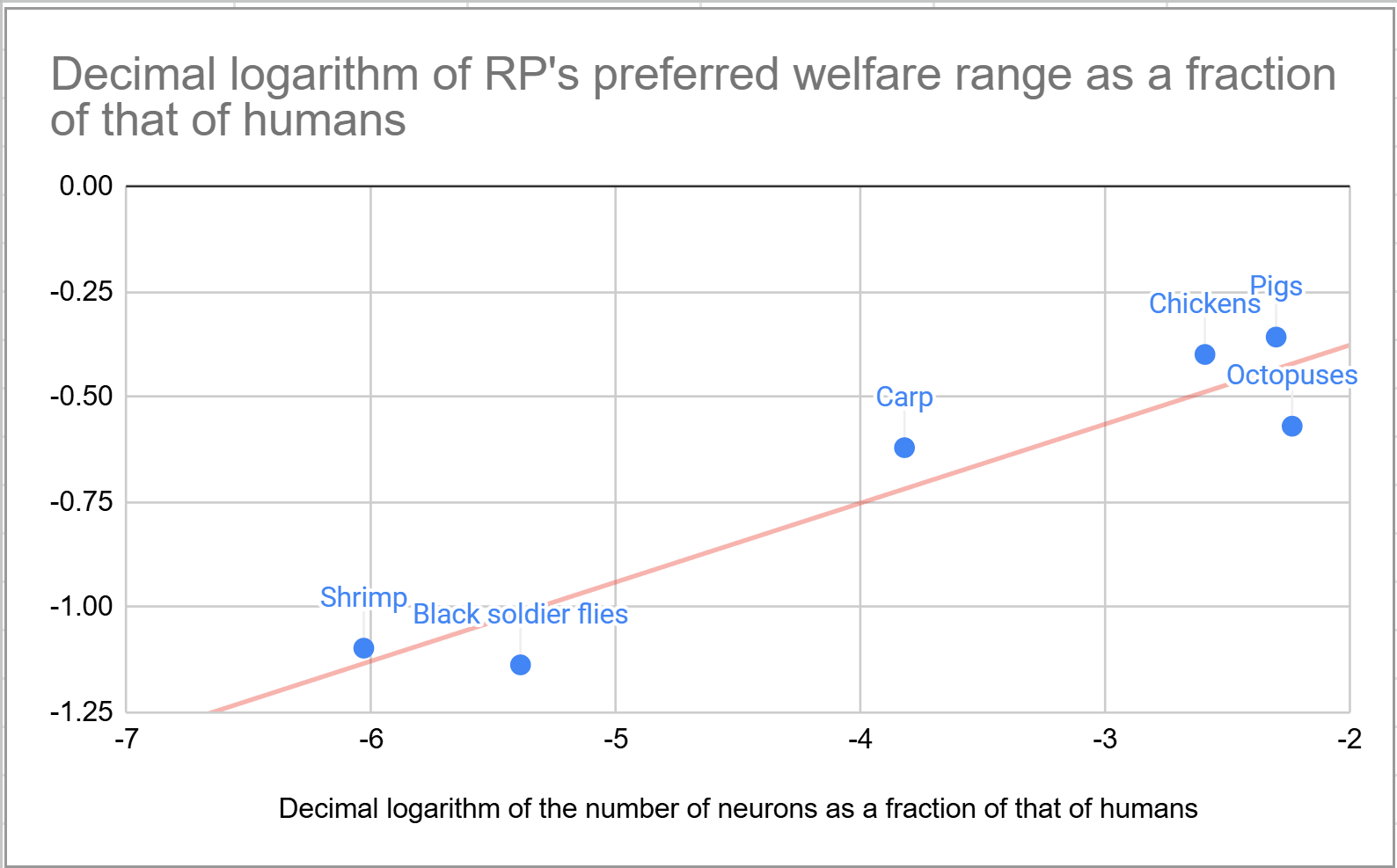
I suppose welfare per animal-year is proportional to the welfare range, the difference between the maximum and minimum welfare per unit time, and that this is a power law of the number of neurons. In particular, I use welfare ranges as a fraction of that of humans equal to “number of neurons as a fraction of that of humans”^“exponent of the number of neurons”, with the exponent ranging from 0 to 2, as I didbefore. For an exponent of:
0, all animals have the welfare range of humans.
0.188:
The welfare ranges are pretty similar to the estimates in Bob Fischer’s book about comparing animal welfare across species, which contains what Rethink Priorities (RP) stands behind now. An exponent of 0.188 explains78.6 % of their variance.
The number of neurons has to become 209 k (= 10^(1/0.188)) times as large for the welfare range to become 10 times as large.
0.5, corresponding to my best guesses for the welfare ranges, the number of neurons has to become 100 (= 10^(1/0.5)) times as large for the welfare range to become 10 times as large.
1, the welfare ranges are proportional to the number of neurons.
2, the number of neurons has to become 3.16 (= 10^(1/2)) times as large for the welfare range to become 10 times as large.
I estimate food consumption (excluding dairy and soy milk):
Increases the living time of directly affected animals by 2.24 (pork) to 9.92 k (shrimp) animal-day/food-kg.
Decreases the living time of soil animals by 11.4 M (shrimp) to 1.39 billion (beef) animal-year/food-kg.
Decreases the living time of soil animals by 418 k (shrimp) to 164 billion (beef) times as much as it increases the living time of the directly affected animals.
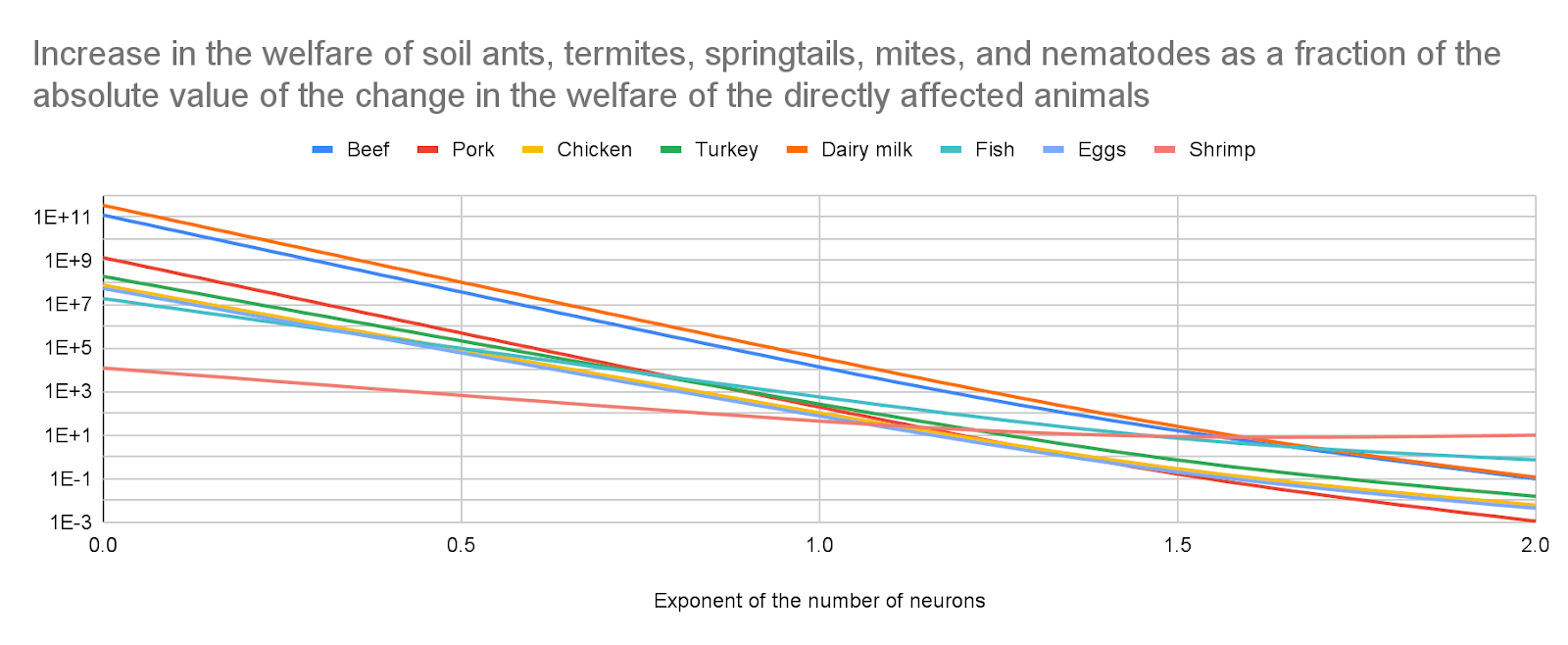
I believe effects on soil animals are much larger than those on the directly affected animals. I am confident the exponent of the number of neurons is the parameter which affects the ratio between the effects on soil animals and directly affected animals the most by far, and effects on soil animals dominate at least for values of the exponent up to 1, which are the ones I consider plausible. I get the following increase in the welfare of soil ants, termites, springtails, mites, and nematodes as a fraction of the absolute value of the change in the welfare of the directly affected animals (a value over 1 implies the effects on soil animals are larger than those on the directly affected animals). For an exponent of the number of neurons of (the lower and upper bound respect shrimp and dairy milk):
0.19, 3.95 k to 15.8 billion.
0.5, 664 to 104 M.
1, 44.1 to 35.9 k.
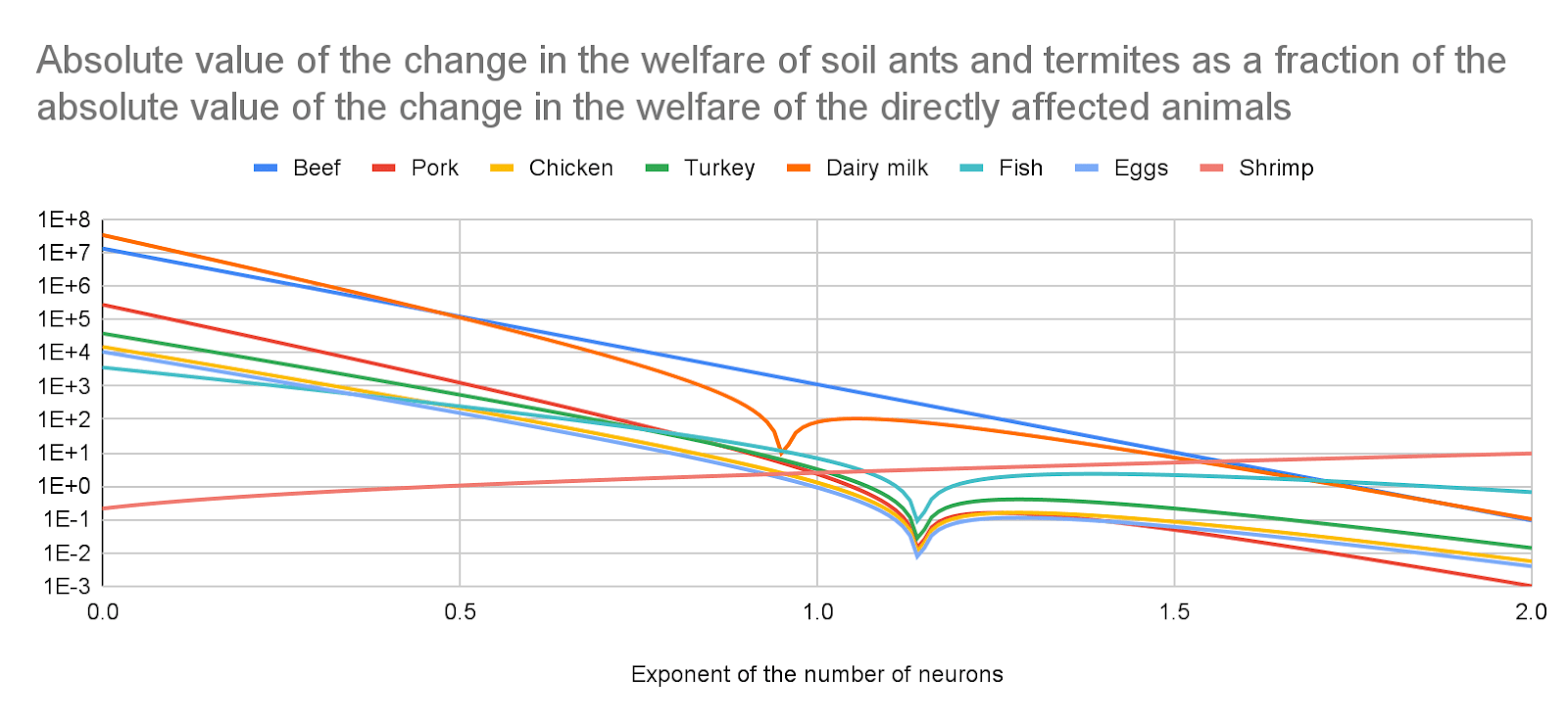
For all the animal-based foods I analysed besides shrimp, I estimate effects on soil animals would still be much larger than those on the target beneficiaries for a welfare per animal-year of exactly 0 for animals with fewer neurons than those considered in Bob’s book, and an exponent of the number of neurons of 0.19 which explains very well its estimates. I calculate soil ants and termites have 2.91 and 1.16 times as many neurons as shrimp, so effects on them would still be relevant. I get the following absolute value of the change in the welfare of soil ants and termites as a fraction of the absolute value of the change in the welfare of the directly affected animals for an exponent of 0.19:
For the animal-based foods I analysed besides shrimp, 1.35 k (fish) to 4.06 M (dairy milk).
For shrimp, 47.4 %.
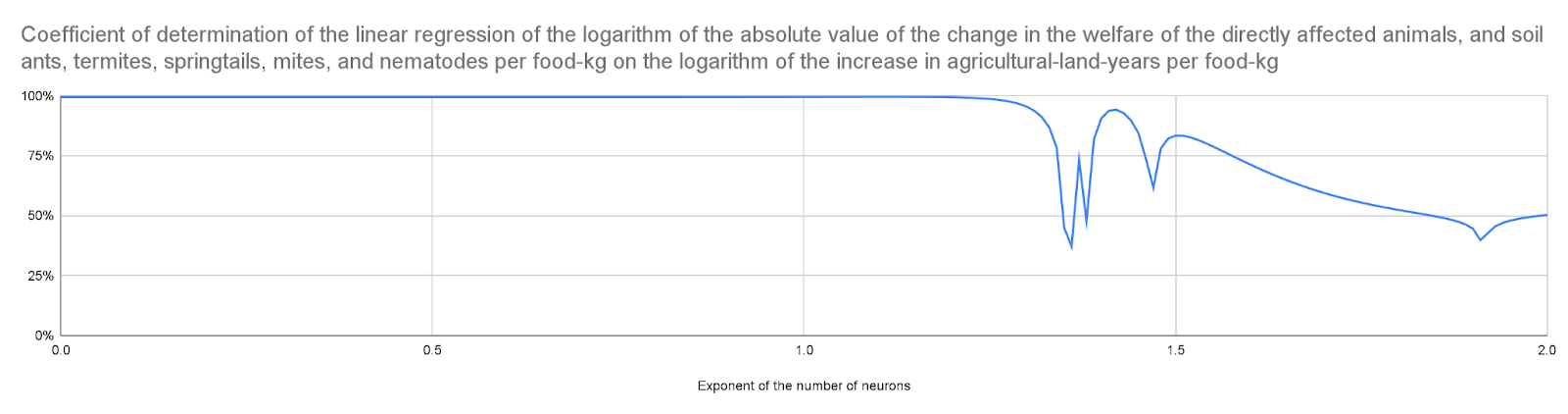
The logarithm of the increase in agricultural-land-years per food-kg explains over 90 % of the variance in the logarithm of the absolute value of the change in the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes per food-kg for an exponent of the number of neurons up to 1.43. In other words, one can predict this absolute value of the change in welfare per food-kg from the increase in agricultural-land-years per food-kg alone for those exponents. This is because the effects on soil animals are much larger than those on the directly affected animals in this case, and the increase in the welfare of soil animals per unit area is similar.
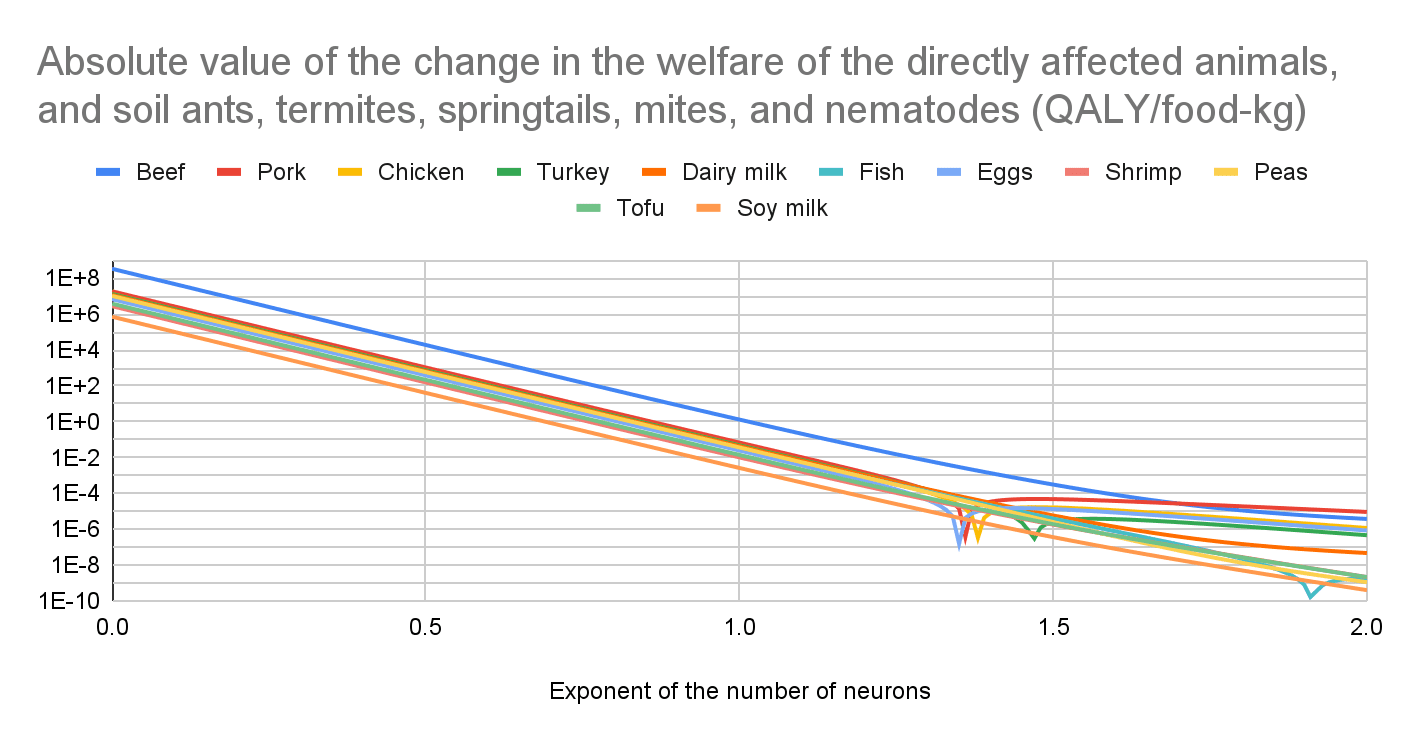
I conclude producing beef increases the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes (considered all together) much more than the other foods for any exponent of the number of neurons.
I continue to recommend changes in food consumption which increase agricultural land. I estimate the m2-years of agricultural land per food-kg almost perfectly explain the increase in the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes per food-kg for my preferred exponent of the number of neurons of 0.5 (see last 2 graphs). In particular, I recommend increasing the consumption of beef, which requires way more agricultural land than the other foods I analysed. Nevertheless, I recommend funding the Centre for Exploratory Altruism Research’s (CEARCH’s) High Impact Philanthropy Fund (HIPF) over that. I estimated buying beef is 3.72 % as cost-effective as funding HIPF, and that this decreases 5.07 billion soil-animal-years per $.
I recommend investigating whether soil ants and nematodes have positive or negative lives, starting with soil nematodes. I estimate effects on soil ants account for most of the effects on soil ants, termites, springtails, mites, and nematodes for all the foods I analysed for an exponent of the number of neurons of 1.40 or higher, and that effects on soil nematodes account for most of those effects for all the foods I analysed for an exponent of 1.21 or lower. I supposed soil nematodes have negative lives, and therefore concluded that increasing agricultural land increases welfare for my preferred exponent of 0.5. Nonetheless, I can easily see them having positive lives, and would conclude in this case that increasing agricultural land decreases welfare.
Paving all the Earth’s land would dramatically decrease the population of soil animals, but is considered bad by many, who therefore argue against decreasing the population of soil animals. However, I am not arguing for paving all the Earth’s land. I am arguing for, by increasing cost-effectiveness, changes in food consumption which increase agricultural land, the most cost-effective global health interventions, and targeted research on whether soil animals have positive or negative lives. I think advocating for paving the whole Earth is much less cost-effective than for the most cost-effective global health interventions.
Furthermore, one could support decreasing the number of soil animals via changing food consumption, and funding global health interventions while strongly believing there should be some wild areas. After millennia of agricultural expansion, the world has passed peak agricultural land. So pursuing interventions increasing agricultural land would only slow down the growth of wild areas.
Methods
I estimate the change in the welfare of animals affected by the production of beef, pork, chicken, turkey, dairy milk, fish, eggs, shrimp, peas, tofu, and soy milk. I analyse directly affected animals, and soil ants, termites, springtails, mites, and nematodes impacted by changes in land use.
I suppose welfare per animal-year is proportional to the welfare range, the difference between the maximum and minimum welfare per unit time, and that this is a power law of the number of neurons. In particular, I use welfare ranges as a fraction of that of humans equal to “number of neurons as a fraction of that of humans”^“exponent of the number of neurons”, with the exponent ranging from 0 to 2, as I didbefore. For an exponent of:
0, all animals have the welfare range of humans.
0.188:
The welfare ranges are pretty similar to the estimates in Bob’s book about comparing animal welfare across species, which contains what RP stands behind now. An exponent of 0.188 explains78.6 % of their variance.
The number of neurons has to become 209 k (= 10^(1/0.188)) times as large for the welfare range to become 10 times as large.
0.5:
I get my best guesses for the welfare ranges.
The number of neurons has to become 100 (= 10^(1/0.5)) times as large for the welfare range to become 10 times as large.
1:
The welfare ranges are proportional to the number of neurons.
The number of neurons has to become 10 times as large for the welfare range to become 10 times as large.
2, the number of neurons has to become 3.16 (= 10^(1/2)) times as large for the welfare range to become 10 times as large.
RP’s moral weight project included a report by Adam Shriver concluding “there is no straightforward empirical evidence or compelling conceptual arguments indicating that relative differences in neuron counts within or between species reliably predicts welfare relevant functional capacities”. I guess there are other factors besides the number of neurons that influence the welfare range. However, an exponent of 0.188 explains 78.6 % of the variance of the estimates in Bob’s book. I get this exponent from the slope of the belowlinear regression with null intercept of the logarithm of RP’s preferred welfare range as a fraction of that of humans on the logarithm of the number of neurons as a fraction of that of humans. I rely on a simple formula for the welfare range to decrease noise, and easily obtain estimates for animals not covered in the book to explore implications for cause prioritisation.
My formula for the welfare range as a fraction of that of humans implies a welfare range of 0 for organisms without neurons, which I think is an underestimate, as I am not certain they have a constant welfare per unit time as a result of not having neurons. Furthermore, I speculate effects on microorganisms, which do not have neurons, are much larger than those on soil animals, although positively correlated.
I calculate the decrease in the welfare of the directly affected animals per food-kg by multiplying my pastestimates by my updated welfare range of the directly affected animals as a fraction of that I used to obtain them.
I suppose the welfare per animal-year of soil ants/termites/springtails/mites/nematodes is -25 % that of fully happy soil ants/termites/springtails/mites/nematodes. I assume this holds for all biomes, but I guess there is variation in reality. My best guess is that soil animals have negative lives. I am very uncertain, but my assumption of negative lives is quite typical. Karolina Sarek, Joey Savoie, and David Moss estimated-0.42 for the “wild bug” in 2018, which is more negative than what I assumed.
I get the number of soil ants, termites, springtails, and mites per unit area for 10 biomes using the means in Table S4 of Rosenberg et al. (2023). I determine the number of soil nematodes per unit area by multiplying the number of soil arthropods from this table by 48.9, which is my estimate for the ratio between the number of soil nematodes and soil arthropods globally.
I set the animal-years of directly affected animals per food-kg of animal-based foods to estimatesfromFaunalytics for the United States (US) for the living time of farmed and wild animals, including farmed animals which die before slaughter, and 1 animal-day per wild feeder fish, which is supposed to be the time from catch to death.
1E+N means 1*10^N. For example, 1E+2 means 1*10^2 = 100.
Number of soil animals affected
I estimate food consumption (excluding dairy and soy milk):
Increases the living time of directly affected animals by 2.24 (pork) to 9.92 k (shrimp) animal-day/food-kg.
Decreases the living time of soil animals by 11.4 M (shrimp) to 1.39 billion (beef) animal-year/food-kg.
Decreases the living time of soil animals by 418 k (shrimp) to 164 billion (beef) times as much as it increases the living time of the directly affected animals.
Food
Increase in the living time of directly affected animals (animal-day/food-kg)
Initial number of soil animals per m² of the affected land
Final number of soil animals per m² of the affected land
Decrease in the number of soil animals per m²
Decrease in the living time of soil animals (animal-year/food-kg)
Decrease in the living time of soil animals as a fraction of the increase in the living time of directly affected animals
Beef
3.09
5.11E+06
8.62E+05
4.25E+06
1.39E+09
1.64E+11
Pork
2.24
5.81E+06
1.36E+06
4.45E+06
7.73E+07
1.26E+10
Chicken
28.7
5.81E+06
1.36E+06
4.45E+06
5.44E+07
6.92E+08
Turkey
11.5
5.81E+06
1.36E+06
4.45E+06
5.44E+07
1.73E+09
Dairy milk
0.0378
6.38E+06
1.01E+06
5.37E+06
4.80E+07
4.64E+11
Fish
82.1
5.81E+06
1.36E+06
4.45E+06
3.74E+07
1.67E+08
Eggs
28.0
5.81E+06
1.36E+06
4.45E+06
2.79E+07
3.64E+08
Shrimp
9.92E+03
5.18E+06
1.36E+06
3.82E+06
1.14E+07
4.18E+05
Peas
0
7.06E+06
1.36E+06
5.70E+06
4.25E+07
Tofu
0
5.81E+06
1.36E+06
4.45E+06
1.57E+07
Soy milk
0
5.81E+06
1.36E+06
4.45E+06
2.94E+06
Welfare range of the directly affected animals as a fraction of that of humans
The welfare range of the directly affected animals decays faster (with the exponent of the number of neurons) for ones with fewer neurons. The slope of the straight lines below is the logarithm of the number of neurons as a fraction of that of humans.
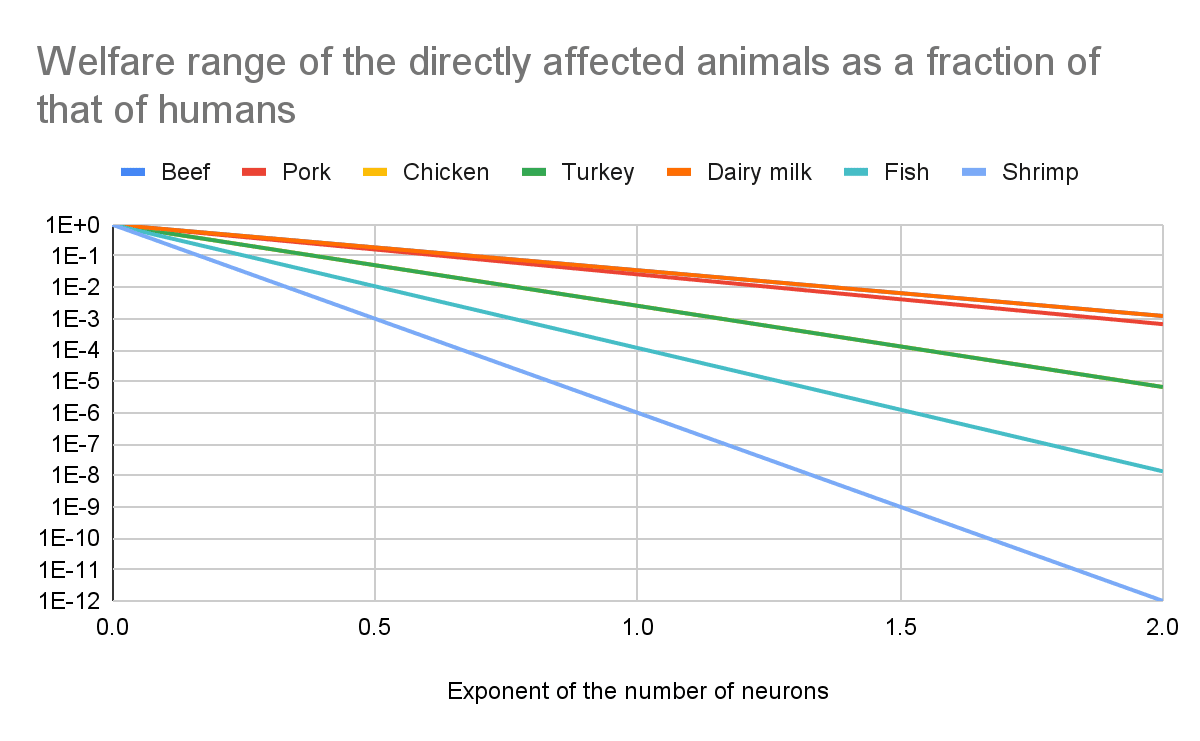
Absolute value of the change in the welfare of the directly affected animals
The lines for beef and dairy milk respect increases in welfare, and all the others represent decreases.
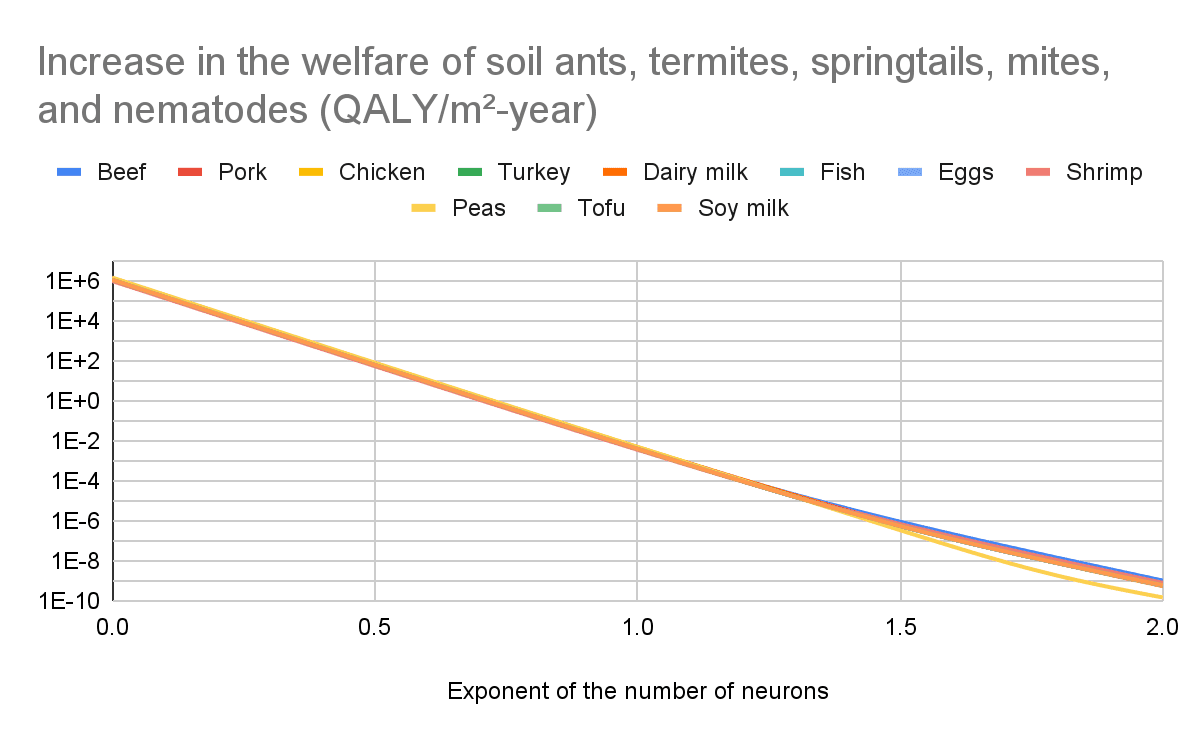
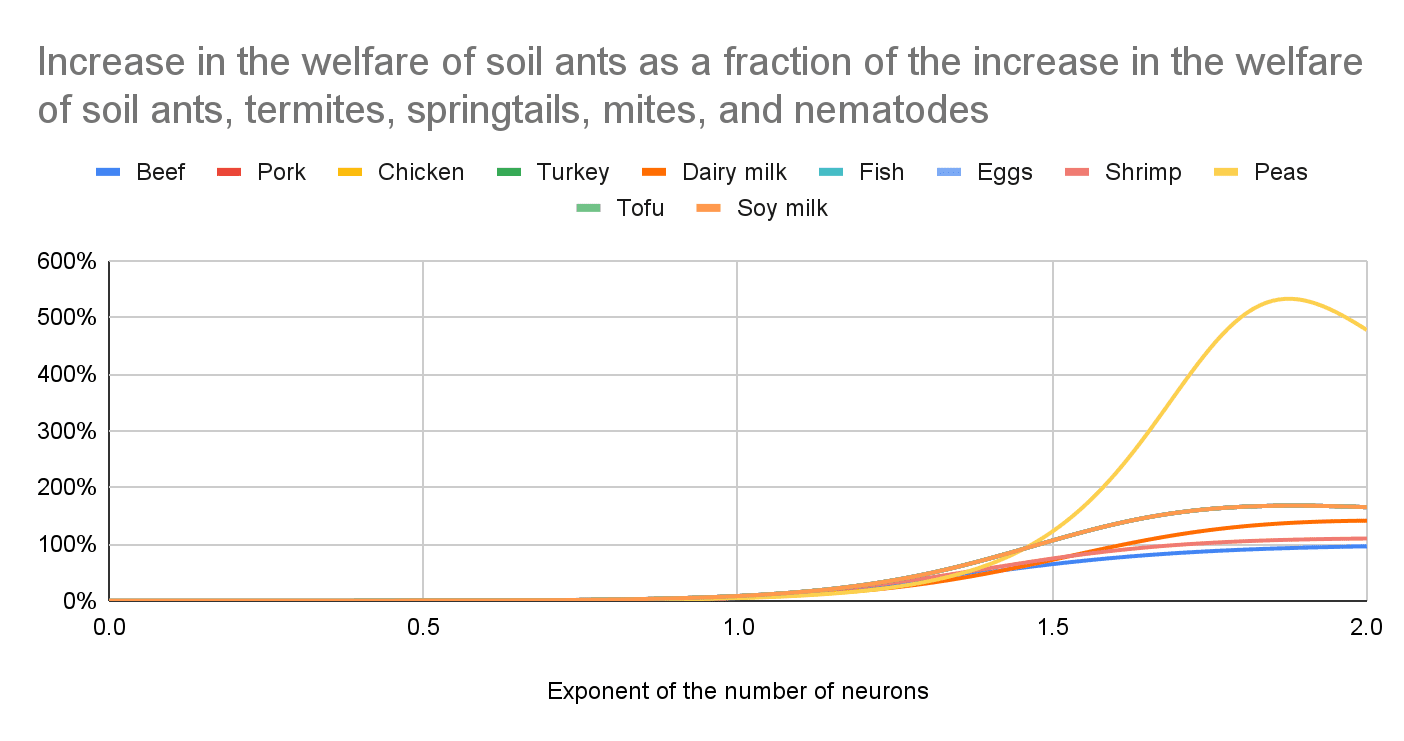
Increase in the welfare of soil ants, termites, springtails, mites, and nematodes
The increase in the welfare of soil soil ants, termites, springtails, mites, and nematodes per unit area is similar for all interventions because Gemini 2.5 guessed the additional agricultural land would replace biomes in approximately the same way. In reality, there is variation even within a single type of intervention.
Increase in the welfare of soil ants as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
The effect on soil ants is the major driver of the effects on soil ants, termites, springtails, mites, and nematodes for a high exponent of the number of neurons because they have the most neurons per individual among those animals.
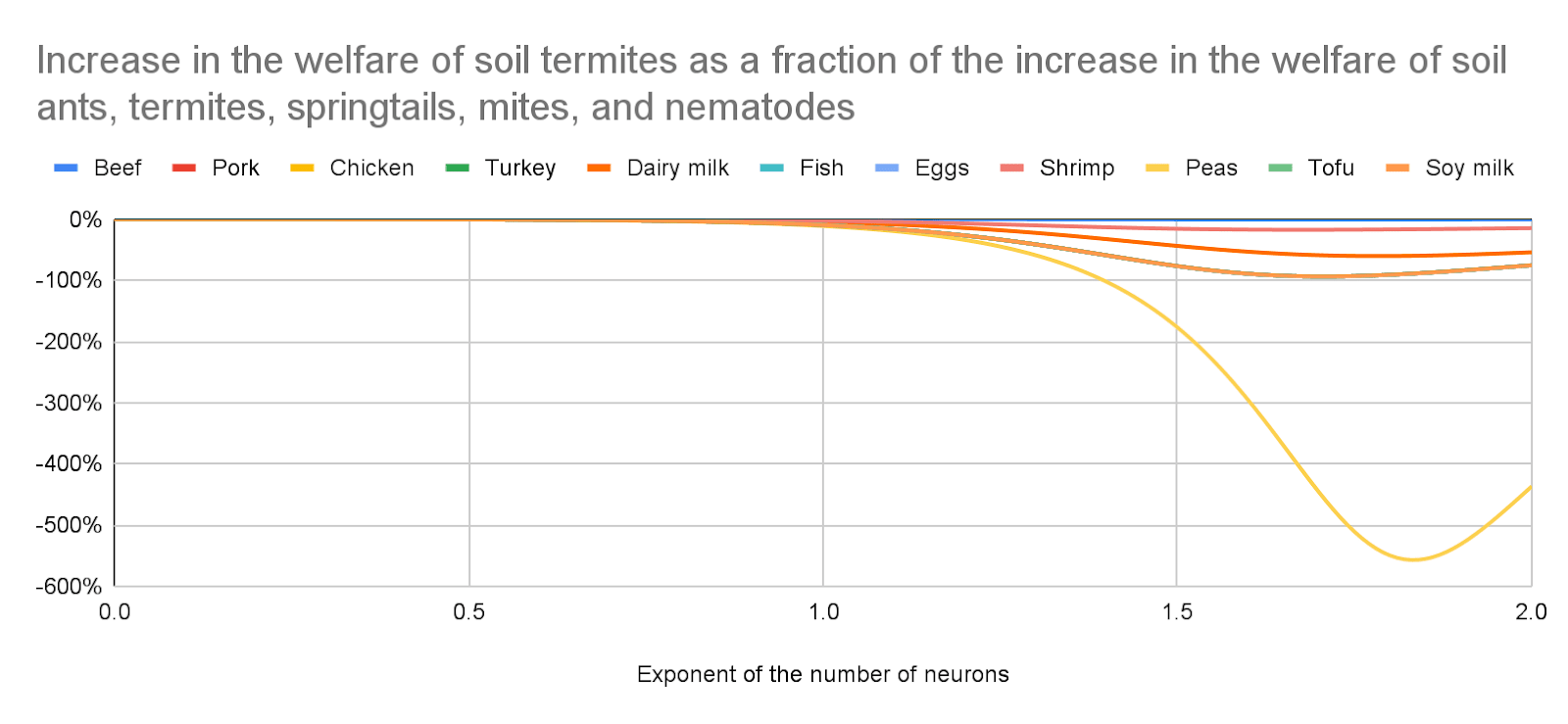
Increase in the welfare of soil termites as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
I infer food production decreases the welfare of soil termites. However, crops and pastures have the least soil ants/springtails/mites/nematodes per unit area besides deserts, and xeric shrublands, which would very hardly be replaced by the additional agricultural land, and effects on soil termites account for a tiny fraction of the effects on soil ants, termites, springtails, mites, and nematodes for an exponent of the number of neurons lower than 1, which I endorse. So I conclude the welfare of those animals considered together would still decrease for land use changes different from the ones guessed by Gemini.
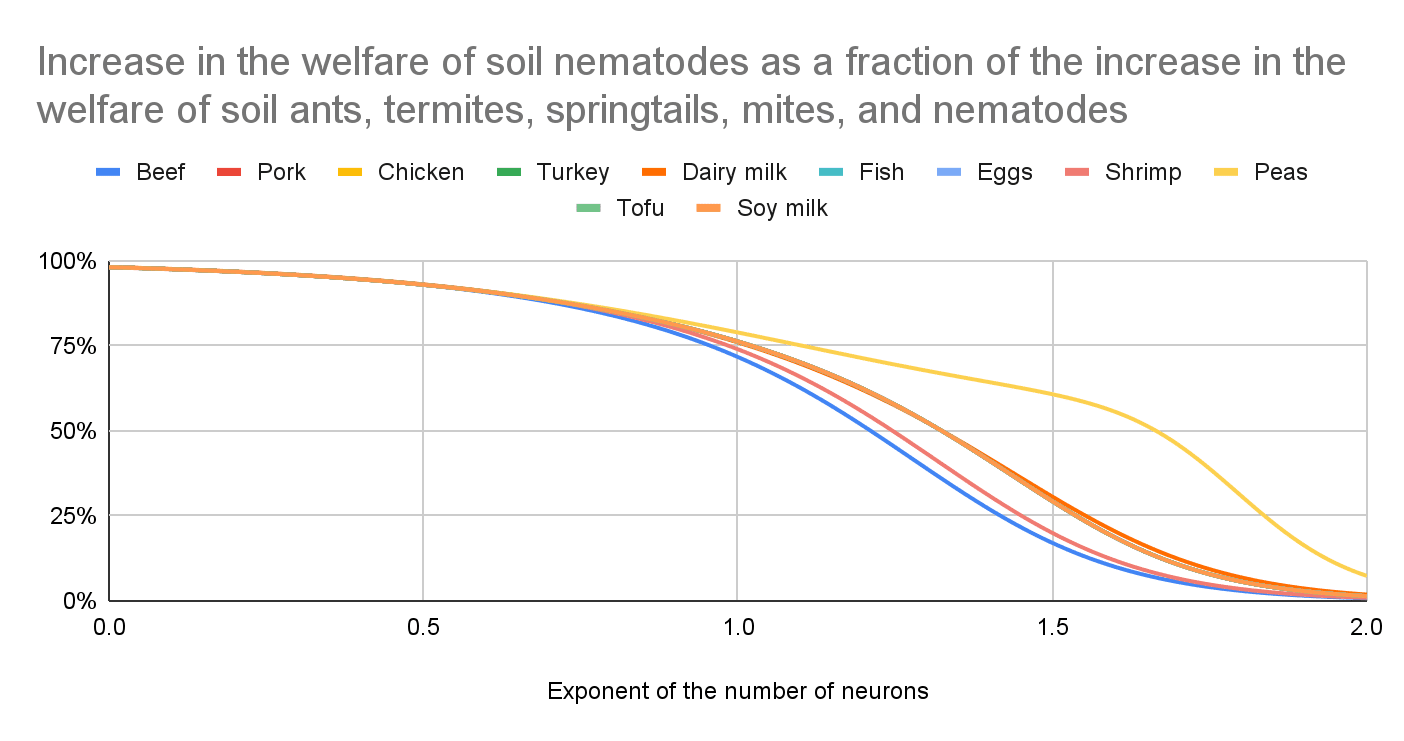
Increase in the welfare of soil nematodes as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
The effect on soil nematodes is the major driver of the effects on soil ants, termites, springtails, mites, and nematodes for a low exponent of the number of neurons because they have the least neurons per individual among those animals.
Increase in the welfare of soil ants, termites, springtails, mites, and nematodes
There is some variation in the increase in the welfare of soil ants, termites, springtails, mites, and nematodes per $ across foods. Yet, there is way more variation with the exponent of the number of neurons within a single food.
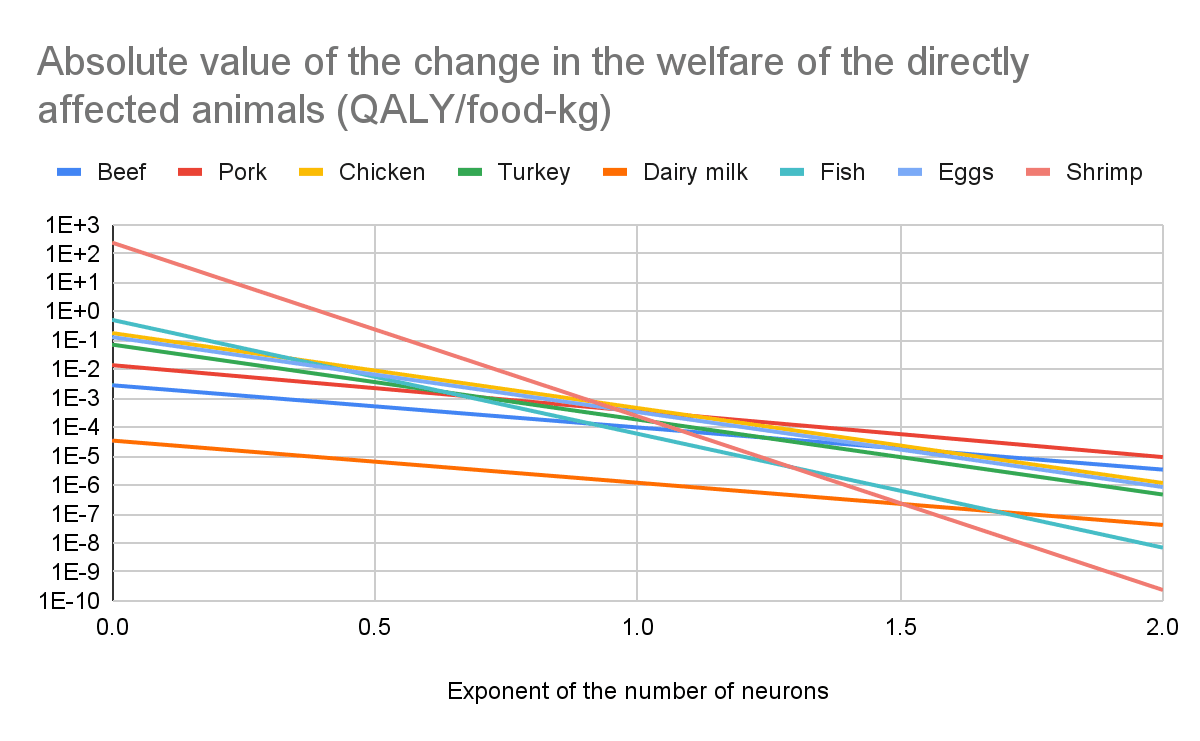
Increase in the welfare of soil ants, termites, springtails, mites, and nematodes as a fraction of the absolute value of the change in the welfare of the directly affected animals
I believe effects on soil animals are much larger than those on the directly affected animals. I am confident the exponent of the number of neurons is the parameter which affects the ratio between the effects on soil animals and directly affected animals the most by far, and effects on soil animals dominate at least for values of the exponent up to 1, which are the ones I consider plausible. I get the following increase in the welfare of soil ants, termites, springtails, mites, and nematodes as a fraction of the absolute value of the change in the welfare of the directly affected animals (a value over 1 implies the effects on soil animals are larger than those on the directly affected animals). For an exponent of the number of neurons of (the lower and upper bound respect shrimp and dairy milk):
0.19, 3.95 k to 15.8 billion.
0.5, 664 to 104 M.
1, 44.1 to 35.9 k.
Absolute value of the change in the welfare of soil ants and termites as a fraction of the absolute value of the change in the welfare of the directly affected animals
For all the animal-based foods I analysed besides shrimp, I estimate effects on soil animals would still be much larger than those on the target beneficiaries for a welfare per animal-year of exactly 0 for animals with fewer neurons than those considered in Bob’s book, and an exponent of the number of neurons of 0.19 which explains very well its estimates (an exponent of 0.188 explains 78.6 % of their variance). I calculate soil ants and termites have 2.91 (= 250*10^3/(86*10^3)) and 1.16 (= 100*10^3/(86*10^3)) times as many neurons as shrimp, so effects on them would still be relevant. I get the following absolute value of the change in the welfare of soil ants and termites as a fraction of the absolute value of the change in the welfare of the directly affected animals for an exponent of 0.19:
For the animal-based foods I analysed besides shrimp, 1.35 k (fish) to 4.06 M (dairy milk).
For shrimp, 47.4 %.
The production of beef and shrimp increases the welfare of soil ants and termites for any exponent. The other animal-based foods decrease it before the sharp points below, and increase it afterwards.
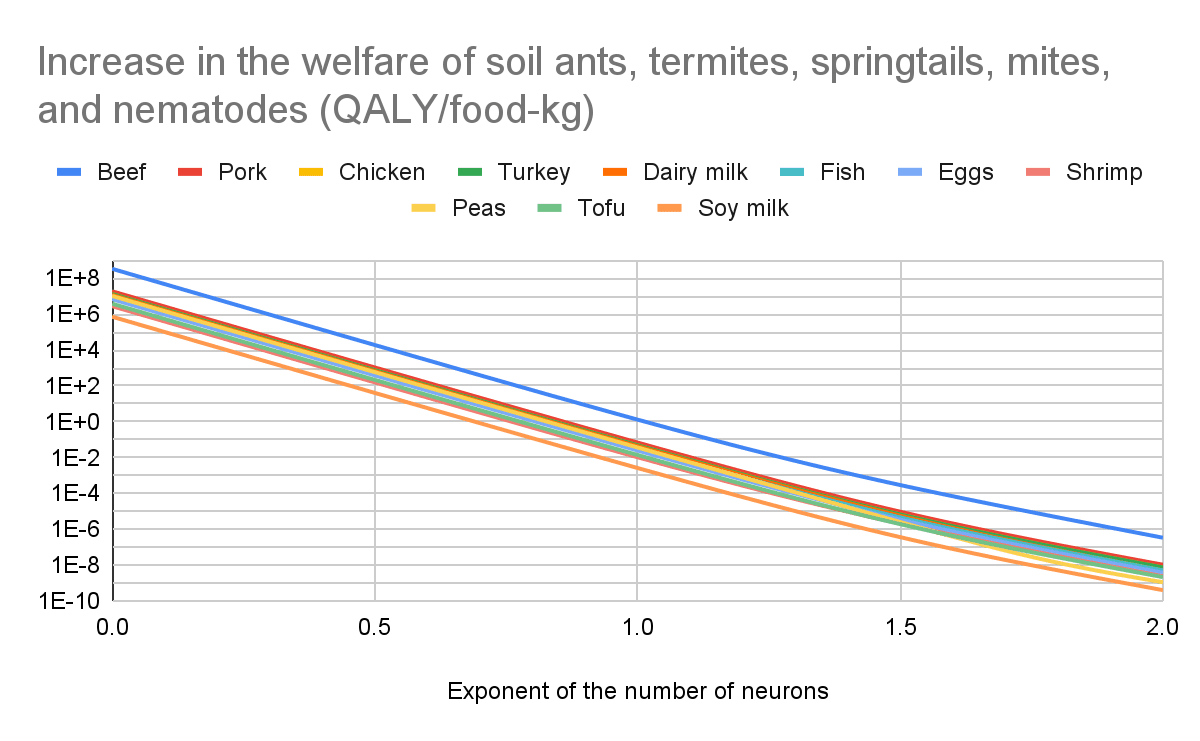
Increase in the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes
I conclude producing beef increases the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes (considered all together) much more than the other foods for any exponent of the number of neurons. The production of beef, dairy milk, peas, tofu, and soy milk increases their welfare for any exponent. The production of pork, chicken, turkey, fish, and eggs increases it before the sharp points below, and decreases it afterwards. The minimum exponent to decrease their welfare is 1.36 for pork, 1.39 for chicken, 1.47 for turkey, 1.92 for fish, 1.35 for eggs, and higher than 2 for shrimp.
Coefficient of determination of the linear regression of the logarithm of the absolute value of the change in the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes per food-kg on the logarithm of the increase in agricultural-land-years per food-kg
The logarithm of the increase in agricultural-land-years per food-kg explains over 90 % of the variance in the logarithm of the absolute value of the change in the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes per food-kg for an exponent of the number of neurons up to 1.43. In other words, one can predict this absolute value of the change in welfare per food-kg from the increase in agricultural-land-years per food-kg alone for those exponents. This is because the effects on soil animals are much larger than those on the directly affected animals in this case, and the increase in the welfare of soil animals per unit area is similar.
Results for my preferred welfare ranges
Below are the results for my preferred welfare ranges respecting an exponent of the number of neurons of 0.5. My exponent is significantly higher than the value of 0.188 which I estimate explains 78.6 % of the variance in RP’s preferred estimates. So my exponent implies the welfare range increases much closer to linearly with the number of neurons, although still significantly sublinearly.
There is variation in land use changes within each type of food, and therefore increasing the production of specific subtypes of food matters. For example, increasing the production of some chicken may increase welfare more than increasing the production of random pork.
Food
Beef
Pork
Chicken
Turkey
Dairy milk
Fish
Eggs
Shrimp
Peas
Tofu
Soy milk
Increase in agricultural land (m²-year/food-kg)
326
17.4
12.2
12.2
8.95
8.41
6.27
2.97
7.46
3.52
0.660
Decrease in the living time of soil animals (animal-year/food-kg)
1.39E+09
7.73E+07
5.44E+07
5.44E+07
4.80E+07
3.74E+07
2.79E+07
1.14E+07
4.25E+07
1.57E+07
2.94E+06
Exponent of the number of neurons regarding my preferred welfare range
0.500
0.500
0.500
0.500
0.500
0.500
0.500
0.500
0.500
0.500
0.500
Welfare range of the directly affected animals as a fraction of that of humans
0.187
0.161
0.0507
0.0507
0.187
0.0108
0.0507
0.00100
Welfare range of the directly affected animals as a fraction of that I have used in the past
36.3%
31.2%
15.3%
15.3%
36.3%
12.1%
15.3%
3.23%
Decrease in the welfare of the directly affected animals (QALY/food-kg)
-5.26E-04
0.00223
0.00905
0.00362
-6.46E-06
0.00550
0.00657
0.238
0
0
0
Increase in the welfare of soil ants (QALY/m²-year)
0.201
0.201
0.201
0.201
0.157
0.201
0.201
0.178
0.143
0.201
0.201
Increase in the welfare of soil termites (QALY/m²-year)
-0.00461
-0.363
-0.363
-0.363
-0.238
-0.363
-0.363
-0.0940
-0.516
-0.363
-0.363
Increase in the welfare of soil springtails (QALY/m²-year)
1.06
2.10
2.10
2.10
2.36
2.10
2.10
0.768
3.35
2.10
2.10
Increase in the welfare of soil mites (QALY/m²-year)
2.97
2.48
2.48
2.48
2.96
2.48
2.48
2.893
2.65
2.48
2.48
Increase in the welfare of soil nematodes (QALY/m²-year)
55.0
57.6
57.6
57.6
69.5
57.6
57.6
49.5
73.8
57.6
57.6
Increase in the welfare of soil ants, termites, springtails, mites, and nematodes (QALY/m²-year)
59.2
62.0
62.0
62.0
74.7
62.0
62.0
53.2
79.4
62.0
62.0
Increase in the welfare of soil ants as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
0.339%
0.325%
0.325%
0.325%
0.211%
0.325%
0.325%
0.334%
0.180%
0.325%
0.325%
Increase in the welfare of soil termites as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
-0.00778%
-0.585%
-0.585%
-0.585%
-0.319%
-0.585%
-0.585%
-0.177%
-0.650%
-0.585%
-0.585%
Increase in the welfare of soil springtails as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
1.80%
3.39%
3.39%
3.39%
3.16%
3.39%
3.39%
1.44%
4.21%
3.39%
3.39%
Increase in the welfare of soil mites as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
5.02%
3.99%
3.99%
3.99%
3.96%
3.99%
3.99%
5.44%
3.34%
3.99%
3.99%
Increase in the welfare of soil nematodes as a fraction of the increase in the welfare of soil ants, termites, springtails, mites, and nematodes
92.9%
92.9%
92.9%
92.9%
93.0%
92.9%
92.9%
93.0%
92.9%
92.9%
92.9%
Increase in the welfare of soil ants, termites, springtails, mites, and nematodes (QALY/food-kg)
1.93E+04
1.08E+03
758
758
668
522
389
158
592
218
40.9
Increase in the welfare of soil ants, termites, springtails, mites, and nematodes as a fraction of the absolute value of the change in the welfare of the directly affected animals
3.67E+07
4.82E+05
8.37E+04
2.09E+05
1.04E+08
9.48E+04
5.92E+04
664
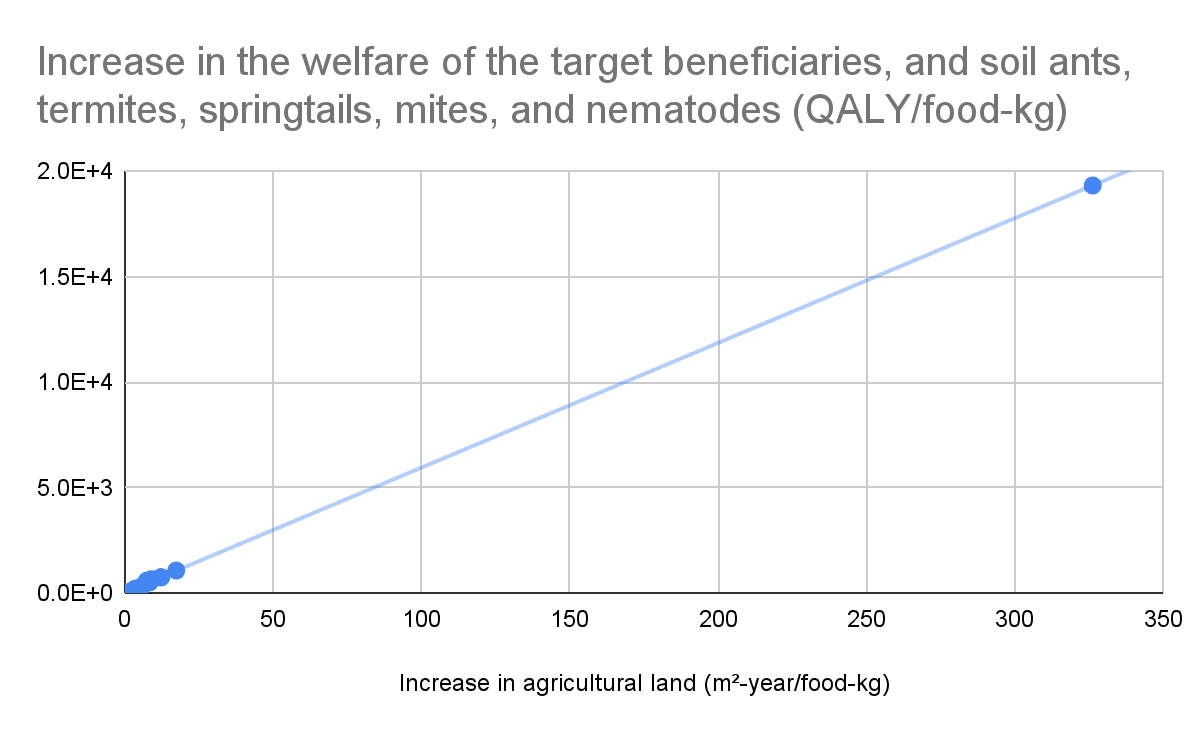
Increase in the welfare of the target beneficiaries, and soil ants, termites, springtails, mites, and nematodes (QALY/food-kg)
1.93E+04
1.08E+03
758
758
668
522
389
158
592
218
40.9
Increase in the welfare of the target beneficiaries, and soil ants, termites, springtails, mites, and nematodes as a fraction of that caused by eating beef
100%
5.57%
3.92%
3.92%
3.46%
2.70%
2.01%
0.817%
3.07%
1.13%
0.212%
For my preferred welfare ranges, the m2-years of agricultural land per food-kg explain 100 % of the variance in the increase in the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes per food-kg.
My recommendations
I continue to recommend changes in food consumption which increase agricultural land. I estimate the m2-years of agricultural land per food-kg almost perfectly explain the increase in the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes per food-kg for my preferred exponent of the number of neurons of 0.5 (see last 2 graphs). In particular, I recommend increasing the consumption of beef, which requires way more agricultural land than the other foods I analysed. Nevertheless, I recommend funding CEARCH’s HIPF over that. I estimated buying beef is 3.72 % as cost-effective as funding HIPF, and that this decreases 5.07 billion soil-animal-years per $.
I recommend investigating whether soil ants and nematodes have positive or negative lives, starting with soil nematodes. I estimate effects on soil ants account for most of the effects on soil ants, termites, springtails, mites, and nematodes for all the foods I analysed for an exponent of the number of neurons of 1.40 or higher, and that effects on soil nematodes account for most of those effects for all the foods I analysed for an exponent of 1.21 or lower. I supposed soil nematodes have negative lives, and therefore concluded that increasing agricultural land increases welfare for my preferred exponent of 0.5. Nonetheless, I can easily see them having positive lives, and would conclude in this case that increasing agricultural land decreases welfare.
Never mind paving all the Earth’s land
Paving all the Earth’s land would dramatically decrease the population of soil animals, but is considered bad by many, who therefore argue against decreasing the population of soil animals. However, I am not arguing for paving all the Earth’s land. I am arguing for, by increasing cost-effectiveness, changes in food consumption which increase agricultural land, the most cost-effective global health interventions, and targeted research on whether soil animals have positive or negative lives. I think advocating for paving the whole Earth is much less cost-effective than for the most cost-effective global health interventions:
I suspect paving decreases soil-animals-years less cost-effectively than funding HIPF:
Concrete costs 425 $/m3 (= (400 + 450)/2). For a concrete depth of 10 cm, 2/3 (= 0.10/0.15) of the 15 cm arguably containing 90 % of the soil animals, paving would cost 42.5 $/m2 (= 425*0.1) ignoring all other costs, or 0.0235 m2/$ (= 1/42.5).
If paving decreased the density of soil animals from my estimate for tropical and subtropical forests of 5.16 M/m2 to 0, it would decrease their density by less than 121 k/$ (= 0.0235*5.16*10^6).
I estimate funding HIPF decreases 5.07 billion soil-animal-year/$. For paving to decrease soil-animal-years as cost-effectively as this, the aforementioned decrease in density would have to last for longer than 983 years (= 5.07*10^9/(5.16*10^6)). Longer because I have not included all the costs of paving. 983 years seems too long.
Advocating for paving wild areas would be much less cost-effective than for the most cost-effective global health interventions even if the cost-effectiveness of funding them was the same.
The cost-effectiveness of advocating for an intervention is the cost-effectiveness of funding it times the money moved to it as a fraction of the money spent advocating for it (fundraising multiplier).
Fundraising for saving lives in low and middle income countries is much easier than for paving wild areas.
Fundraising for paving all the Earth’s land would be even more difficult than for simply more paving of wild areas at the margin.
Furthermore, one could support decreasing the number of soil animals via changing food consumption, and funding global health interventions while strongly believing there should be some wild areas. After millennia of agricultural expansion, the world has passed peak agricultural land. So pursuing interventions increasing agricultural land would only slow down the growth of wild areas.
AI Use Note: Main body text entirely human written. Claude (Opus 4.8) helped develop models of animal life histories in the appendix.
Cross-posted from Good Structures.
Executive Summary
* Animal advocates sometimes make claims like “there are X of this animal...
Adapted from my Substack, Funding Anthropalypse.
Short version: if you want a share of the coming Anthropic and OpenAI windfall - the $37bn+ that could be in play next year - the way in is to become 'legibly excellent', so the evaluators and donors that frontier lab staff already trust point them to yo...
Disclaimer: Although I work on the Groups Team at CEA, I’m writing this in a personal capacity, and this post does not constitute an endorsement by CEA.
Agency - the realisation that you really can just do things.
TL;DR
Biosecurity needs people (of any background) who are agentic and have a high execution velocity and track record....


Hi Vasco. I am new on this forum, so please excuse my lack of knowledge on the specifics of your work over the years. I see that you produce high quantities of seemingly high quality work on undervalued topics such as this one. I myself am very much interested in the sentience and welfare of such “primitive” organisms. Here are a few of my impressions on this post:
Thank you for sharing your work here. I do not expect you to answer thoroughly to every point I made if you do not think your time is worth it - I just felt like sharing a few thoughts on an interesting topic and participating in the forum - but I will gladly read your responses (or anyone's) if you have some.
Great comment and welcome to the forum! looking forward to hearing more of your thoughts :).
Only one small comment which may help understanding @Vasco Grilo🔸 can say if I'm representing him incorrectly. I agree that the uncertainty is so high here that it doesn't make sense to make strong practical recommendations. Vasco though is a mathematical utilitarian in a pretty pure form, so he's seemingly happy to make strong recommendations where there's little evidence and probabilities are close to 50/50. he'll then even change those recommendations immediately after doing some more calculations. I don't really understand how this can work in practice as communities of EA doers obviously can't switch from advocating eating less meat to advocating huge farms on the basis of an extremely uncertain BOTEc. I've made a similar point to you on as few of his posts in the past.
Thanks for the relevant comment, Nick. I strongly upvoted it because I felt it was useful for me to think about this more.
I recommend funding A over B if I think funding A increases welfare more cost-effectively at the margin than funding B (in expectation). However, my numerical estimates of the cost-effectiveness do not integrate all the information I have. If my numerical estimates for the cost-effectiveness of 2 interventions are similar, other factors could easily be decisive.
I agree it would not make sense for a whole community to switch back and forth between 2 interventions with similar positive cost-effectiveness at the margin. The amount of resources moving from the most to the least cost-effective intervention at the margin should tend to 0 as the difference between the marginal cost-effectiveness of the interventions tends to 0.
I also agree it would not make for a single person to be constantly switching back and forth between 2 interventions with similar positive cost-effectiveness at the margin. Spending 1 year to switch to a career path which is 0.1 % more impactful per year, accounting for impact through work and donations, would only make sense if one could work for longer than 1 k years (= 1/10^-3) in the new path. This is way too long, and therefore the change would not be worth it.
However, I think the situation is different in the context of constantly switching back and forth between supporting or not an intervention due to large uncertainty about whether it increases or decreases welfare (in expectation). Constant switching leads to spending resources without achieving nothing, which is worse than supporting positive interventions. However, it is unclear whether achieving nothing is better or worse than supporting an intervention which can easily have a positive or negative cost-effectiveness.
In cases where there is large uncertainty about whether an intervention increases or decreases welfare (in expectation), I believe it is often better to support interventions decreasing that uncertainty. This is a major reason for my top recommendation of decreasing the uncertainty about whether soil nematodes have positive or negative lives, which I consider robustly better than constant switching. As I say in the summary, "I am arguing for, by increasing cost-effectiveness, changes in food consumption which increase agricultural land, the most cost-effective global health interventions, and targeted research on whether soil animals have positive or negative lives".
Thanks for sharing your thoughts, and welcome to the EA Forum, Guillaume!
I recommend decreasing the living time (total animal-years), not lifespan (animal-years per animal), of soil animals via decreasing their density (animals per unit area) given my best guess that they have negative lives.
Right.
I calculate increasing the consumption of beef by 1 kg results in 1.39 billion fewer soil-animal-years, but just in 3.09 more cow-days. In other words, I estimate increasing the consumption of beef decreases the living time of soil animals 164 billion times as much as it increases the living time of cows. I think there is way more suffering in 164 billion soil-animal-years than in 1 cow-year. I calculate increasing the consumption of beef increases the welfare of soil ants, termites, springtails, mites, and nematodes (due to decreasing their living time) 113 k times as much as it changes the welfare of cows for my preferred exponent of the number of neurons of 0.5. The ratio of 113 k is much lower than that of 164 billion because cows have more intense experiences than soil animals. However, increasing beef consumption affects so many more soil-animal-years than cow-years that I still conclude the effects on soil animals clearly dominate.
I do not. I estimate that funding HIPF increases agricultural land 25.0 (= 1.29*10^3/51.6) times as cost-effectively as buying beef. As I say in the summary:
I think the most cost-effective global health interventions decrease the living time of soil animals much more cost-effectively than interventions aiming to improve the conditions of farmed animals. I estimate that cage-free and broiler welfare corporate campaigns decrease soil-animal-years 1.13 % (= 5.77*10^7/(5.09*10^9)) and 6.50 % (= 3.31*10^8/(5.09*10^9)) as cost-effectively as funding HIPF.
It is very unclear whether global warming increases or decreases animal welfare. I think it would be valuable to investigate the effects of global warming on soil animals, but this is not my top priority. For a given spending on a land use change intervention (like buying beef, or funding HIPF), I would be surprised if the effects of GHG emissions on soil animals were larger than the direct effects on them resulting from changing their density (the abundance of soil animals per unit area varies by biome).
In any case, the effects of GHG emissions being larger would be a reason for increasing or decreasing (depending on what would benefit soil animals) GHG emissions as cost-effectively as possible. If decreasing GHG emissions was among the most cost-effective ways of increasing the welfare of soil animals, I would recommend Founders Pledge's Climate Fund (FPCF), not interventions targeting farmed animals.
I do not see how decreasing GHG emissions would be a cost-effective way of fundraising for wild animal welfare. I estimate only 5.03 M 2023-$ were spent on wild animal welfare in 2023. This is a very tiny fraction of the global economy, so decreasing spending in areas outside wild animal welfare would basically have no impact on its funding.
Yes, I guess the probability of soil animals having negative lives is around 55 %. Research on whether soil animals have positive or negative lives would be valuable in either case, and this is a major reason I recommend it over funding HIPF.
I prefer sharing my uncertain views about topics I have thought about over being agnostic. There are no interventions outside research which I am reasonably confident increase welfare in expectation. I think electrically stunning shrimp is one of those which more clearly increases welfare in expectation, but I would say it is still unclear whether it increases or decreases welfare in expectation due to effects on soil animals. For my individual welfare per animal-year proportional to "number of neurons"^0.5, I determined electrically stunning shrimp increases the welfare of these by 0.00144 QALY/shrimp. There are 94.3 shrimps per shrimp-kg. So infer electrically stunning shrimp increases the welfare of these by 0.136 QALY/shrimp-kg (= 0.00144*94.3). For my individual welfare per animal-year proportional to "number of neurons"^0.5, I estimate replacing farmed shrimp with farmed fish changes the welfare of soil ants, termites, springtails, mites, and nematodes by 364 QALY/shrimp-kg (= 522 - 158). So I conclude electrically stunning farmed shrimp changes the welfare of soil animals more than it increases the welfare of shrimps if it results in the replacement of more than 0.0374 % (= 0.136/364) of the consumption of the affected farmed shrimp by farmed fish. I can easily see this happening for even a slight increase in the cost of shrimp. Moreover, there are 3*10^29 soil archaea and bacteria, 613 M (= 3*10^29/(4.89*10^20)) times as many as soil nematodes (the most abundant soil animals), and I would not be surprised if the effects on soil archaea and bacteria were much larger than those on soil animals, which further contributes to the uncertainty about whether electrically stunning shrimp increases or decreases welfare in expectation.
Side note. Personally, I value decreasing 1 h of suffering as much as creating 1 h of happiness if they have the same intensity. However, many people are suffering-focussed, and therefore value averting 1 h of suffering more than creating 1 h of happiness even if they have the same intensity, and do not want to create additional animal lives even if they are positive. For such suffering-focussed people, it matters less whether soil animals have positive or negative lives. I believe there is a pretty strong case for them wanting to increase agricultural land due to this decreasing animal-years a lot.
I do not think there is a contradiction. One could work on decreasing GHG emissions while driving a car to the office to save time, and therefore decrease GHG emissions more. Likewise, one could argue for increasing the consumption of beef while not eating beef to be more productive, and save money to donate more.
I estimate increasing the consumption of beef by 1 kg results in 9.06 fewer human-minutes due to negatively affecting human health, which would be 1.72*10^-5 (= 9.06/60/24/365.25) fewer QALYs assuming only fully happy life is lost (which slightly overestimates the negative effects on humans). At the same time, I calculate increasing the consumption of beef by 1 kg results in 1.39 billion fewer soil-animal-years, and increases the welfare of the directly affected animals, and soil ants, termites, springtails, mites, and nematodes by 19.3 kQALY for my preferred exponent of the number of neurons of 0.5. So I estimate the increase in the welfare of animals is 1.12 billion (= 19.3*10^3/(1.72*10^-5)) times the decrease in the welfare of humans.
Thank you too for engaging with the post!
I think the focus on beef consumption is better explained by flipping the argument: if you are advocating for a reduction in beef consumption, you may be advocating for a substantial increase in animal suffering, and this should give you pause if the reason you are advocating is that you care deeply and desperately about animal suffering.
The general conclusion being that more research is needed in this area for people who care about soil animal welfare to work out whether soil animals live positive or negative lives, and only once this is done work out how this might be tractably actioned to increase or decrease the number of soil animals via land use change.
I'm honestly really hoping that soil animal lives work out net positive at the mite and springtail level, and that nematodes are sufficiently simple as to not possess the kind of conscious experience needed for welfare capacity. But I remain open to the troubling possibility of well-evidenced alternate conclusions, and support Vasco's work to shed light on this area.
Thanks, Kestrel. I would also prefer soil animals to have positive lives, which would mean a much higher global welfare. However, that also lead to counterintuitive conclusions. I would estimate interventions increasing agricultural land, like cage-free and broiler welfare reforms, and saving human lives, would decrease welfare.